Embryo series courtesy of Einhard Schierenberg
Embryo series courtesy of Einhard SchierenbergTable of Contents
Abstract
The C. elegans tubulin family is composed of nine α-, six β-, and one γ-tubulin. Tubulins are highly conserved, functioning as α-β heterodimers that assemble into microtubules. These cylindrical and ubiquitous components of the cytoskeleton are critical for nearly all cellular and developmental processes. C. elegans has provided a model for the study of microtubules in multiple settings including separation of chromosomes, cellular polarity, and neuronal sensation. Tubulins and microtubules interact with a long list of other cellular proteins that regulate tubulin homeostasis, modify microtubule dynamics, and control incorporation into or disassociation of higher-order cellular structures such as spindles or ciliary axonemes. A collection of enzymes modifies tubulins, often at the variable carboxyl-terminal tail, adding another layer of regulation to microtubule structure and function. Genetic and cytological studies in C. elegans have revealed roles for tubulin and its associated proteins in numerous contexts from embryogenesis to adult behavior.
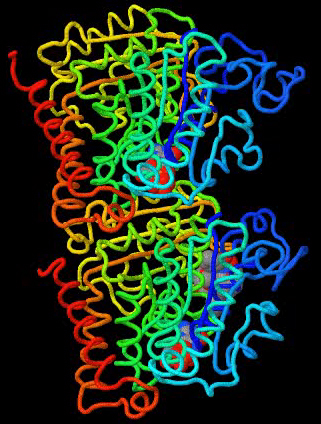
Tubulins are ~450 amino acid globular proteins of the cytoplasm that comprise microtubules (Figure 1). The sequences of both α- and β-tubulins (Figure 2), which form heterodimers, indicate that they are part of a large family of proteins that bind to and hydrolyze guanosine triphosphate (GTP). Tubulin-like proteins appear early in evolution, and they have remained highly conserved in the eukaryotic lineage. Tubulins are related to the FtsZ proteins of eubacteria. Like tubulins, FtsZ proteins polymerize and depolymerize with a GTPase cycle and play a role in cell division (Ingerson-Mahar and Gitai, 2012). Genes for FtsZ homologs are widely distributed in prokaryotic lineages, but closer relatives to eukaryotic tubulins are found in a few archaeal species (Yutin and Koonin, 2012). Amongst eukaryotes, the α- and β-tubulins are nearly identical to each other within their families, as most of the sequence divergence is located in the carboxyl-terminal tails which emanate from the outer surface of the microtubule and can impart specific functions to tubulins, perhaps through interactions with motor proteins and/or other microtubule-associated proteins (Hsu et al., 2014; Nogales et al., 1999; Popodi et al., 2008).
|
Figure 1. Rainbow decorated three-dimensional structure of the tubulin dimer. Colors indicate position in the primary structure as in Figure 2. The amino-terminal GTP binding elements (Rossman fold) span from blue to green and face the interior of the microtubule. Space-filling ligands include GTP bound to both α-tubulin (upper) and β-tubulin (lower), and paclitaxel (taxol) bound to β-tubulin. The external surface of the microtubule is composed of the yellow through red secondary structures. Carboxyl-terminal tails are not part of the crystal structure of the dimer. Animation was created using FirstGlance in Jmol (http://firstglance.jmol.org) and the PDB identification code 1TUB (Nogales et al., 1999).
|
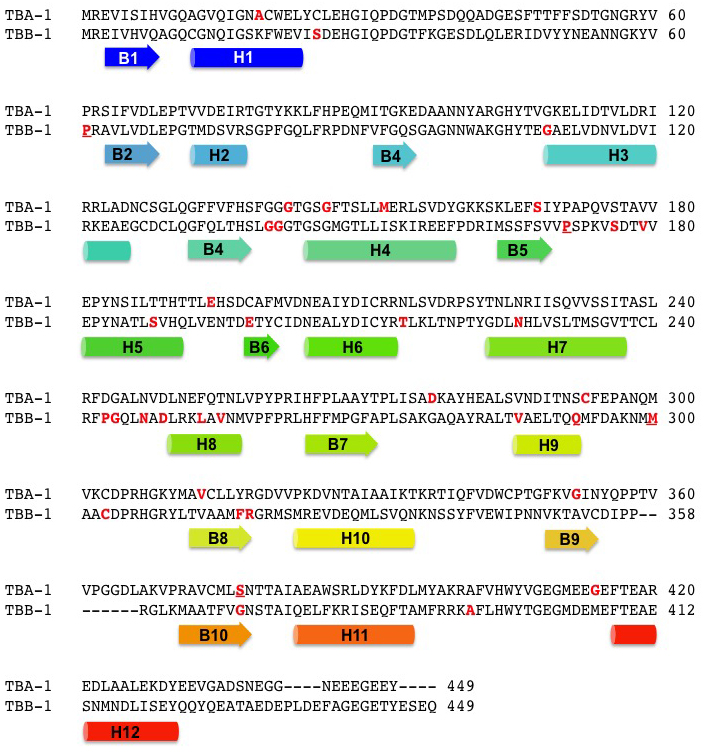
Figure 2. A pairwise alignment of TBA-1 and TBB-1 with dominant alleles and secondary structure. Genetically defined dominant, semi-dominant, and/or gain of function alleles are shown in red and underlined if they have been independently discovered more than once (Table 1). Colored α-helices (cylinders) and β-ribbons (arrows) as in Figure 1 are shown below the primary structure.
Microtubules are semi-rigid, cylindrical cytoskeletal polymers composed of repeating units of the α- and β-tubulin heterodimer. Heterodimers, which are formed soon after synthesis of monomers, associate head to tail into protofilaments. Kinetic and structural data suggest that the further assembly of heterodimers and short protofilaments into microtubules is explained by a lattice model. This model suggests that incorporation into a microtubule straightens soluble, curved heterodimers, while nucleotide status tunes the strength of contacts within the lattice of dimers. Hydrolysis of GTP to GDP in the exchangeable site (E-site) of β-tubulin within a microtubule causes compaction and other conformational changes in both subunits of the dimer that weaken the contacts holding the lattice together, making the microtubule unstable (Rice et al. 2008; Alushin et al., 2014). The structure and nucleotide hydrolysis capacity of tubulin imparts microtubules with dynamic instability, an intrinsic property of self-assembly and disassembly (Mitchison and Kirschner, 1984).
In vivo, microtubules are commonly nucleated by the γ-tubulin ring complex and other components of centrosomes (Teixido-Travesa et al., 2012), which pattern protofilaments into the helical pitch of a microtubule. Nematode microtubules are 11 protofilaments in most cells, while typical eukaryotic microtubules are 13 protofilaments. Touch receptor cells in nematodes have 15 protofilament microtubules (Chalfie and Thomson, 1982). Microtubules are polar polymers, with assembly and disassembly occurring preferentially at the plus end. Similar activities take place at the minus end, although at a slower rate. Polymer polarity is a hallmark of microtubules (and actin), and it often establishes and/or maintains cellular polarity (e.g., see Dubey et al. 2015).
Mutations in genes encoding tubulins in C. elegans have been discovered in screens for a variety of phenotypes. The forward genetic approach has produced a large collection of alleles for which there is a rich and growing collection of phenotypic information. This allows for understanding the cellular role of tubulin in many different contexts (see Section 2). In addition, because the C. elegans genome has a fairly large family of tubulin genes (with many being non-essential), dominant mutations in tubulin have been isolated and propagated. Mapping of these alleles to the structure has provided insight into the function of tubulin, especially in the case of genetically explored and molecularly identified point mutations (Figure 2 and Table 1, Section 6.1).
Tubulin can be divided into three domains. Domain one spans the first 200 amino acids, and it includes the Rossman fold and nucleotide-binding portion that faces the lumen of the microtubule. Dominant mutations have been found in the glycine-rich stretch (GGGTG[T/S]G, sometimes called the tubulin signature motif) in between B4 and H4 of both α and β-tubulin (Figure 1 and Figure 2). Dominant mutations of the first of a conserved set of prolines (aa170) that reside in the pocket around the purine base of GTP have been found twice, both in β-tubulin. Other dominant mutations are scattered in the amino-terminal half of both tubulins, often in between or near the ends of α helices or β ribbons. Two intragenic revertants of a dominant mutation in the conserved S169 have also been identified.
The second domain (sometimes called the activation domain) spans the remaining conserved amino acids up to the divergent C-terminal tails (CTTs, Figure 3). As noted by Erickson, (1998), dominant mutations near or in the highly conserved G244, N247, and D249 of the ‘synergy loop’ have only been found in β-tubulin, typically in mec-7, although the nearby L253 in H8 is altered in an allele of tbb-4. This collection of alleles is curious as the amino acids they change in β-tubulin form part of the binding pocket for the non-exchangeable GTP nucleotide in the dimer-dimer interface that is mostly bound by α-tubulin (Figure 1 and Figure 2). More exploration will reveal whether this is specific to these worm tubulins or telling something broader about dimer formation and the non-exchangeable GTP. A mutation of the conserved methionine at position 300 has been independently isolated twice (both in mec-7), and the serine at position 377 is mutated to phenylalanine in two independent alleles of tba-1. Negative charges around a glycine (aa416 in mec-12, aa414 in Figure 2) near H12 modulate the interaction between microtubules and motor proteins such as dynein (Hsu et al., 2014). As is the case in the amino terminal portion of tubulin, many of the dominant mutations are in between or near the ends of the α-helices and/or β-ribbons. None have yet been found in a CTT.
|
Figure 3. Multiple alignment of α-tubulin and β-tubulin C-termini. An alignment of the C-terminal tails (CTTs) of the nine α-tubulins and six β-tubulins. The sequence shown starts at the end of helix 12 (Figure 2). Tubulins within each family are nearly identical to each other throughout the rest of their structure. The axonemal motif in TBB-4 is shown in bold. For an alignment of complete sequences of all the tubulins in C. elegans, see Gogonea et al., 1999.
The C. elegans tubulin superfamily consists of nine α-tubulins, six β-tubulins, and a single γ-tubulin (Table 2, Section 6.2). The presence of multiple, closely related α- and β-tubulin paralogs (also referred to as isoforms or isotypes) in eukaryotic genomes has historically suggested three hypotheses, which can also be applied to other families of paralogous proteins. First, multiple genes may be present to ensure a stable and redundant pool of tubulin; subtle sequence changes are simply due to post-duplication genetic drift. Second, particular tubulins may be required for specific environmental conditions and are expressed in response to these conditions. Third, the subtle sequence changes of the monomers impart functional specificity to tubulin dimers, which can optimize the function of the microtubule cytoskeleton for its role in a particular cell type or developmental context (Cleveland, 1987; Wade, 2007; Wilson and Borisy, 1997). These explanations are not mutually exclusive as genetic and cytological evidence support redundancy and specificity among the members of the tubulin superfamily in C. elegans and other metazoans.
Worm tubulins have been studied for thirty years in multiple cellular and developmental contexts. C. elegans research made early contributions to establishing a linkage between mechanosensation and microtubule structure/function (Chalfie and Thomson, 1982; Fukushige et al., 1999). These pioneering genetic studies mostly supported the notion that certain tubulins play specific biological roles. Further support for this came from the early observation that certain monoclonal tubulin antibodies labeled specific subsets of neuronal cells (Siddiqui et al., 1989). More recently, non-motile primary cilia at the tip of the dendrite of many sensory neurons have provided a model to study intraflagellar transport processes (see WormBook chapter The sensory cilia of Caenorhabditis elegans; Müller et al., 2011), and they give another example of the use of specific tubulins to optimize behavioral responses (Hao et al., 2011; Hurd et al., 2010).
As a core component of the metazoan cytoskeletal system, microtubules composed of the less diverged, more broadly expressed, and likely more redundant tubulins play numerous and essential roles at all phases of the cell and life cycle. The early C. elegans embryo has become a model to study particular processes such as the assembly and function of the spindle (see Figure 4 and WormBook chapter Cell division; Müller-Reichert et al., 2010; Lu and Mains, 2005; Lu et al., 2004; Cheeranbathur et al., 2013), microtubule dynamics (Bajaj and Srayko, 2013), or the accumulation of spindle proteins after nuclear envelope breakdown (Hayashi et al., 2012). In addition, the early embryo is also a venue to study the role of microtubules in the developmental processes of establishing polarity through asymmetric cell divisions and axis formation, although it is disputed whether centrosomes or the microtubules that emanate from them are the initial symmetry breaking entities (for a history of the evidence both ways, see Wallenfang and Seydoux, 2000; Cowan and Hyman, 2004; Tsai and Ahringer, 2007; Motegi et al. 2011; Bienkowska and Cowan, 2012).
|
Figure 4. Translational fusions to the promoter of pie-1 allow for live visualization of tubulins and other components of mitosis in embryonic blastomeres. Strain SA250 contains GFP::TBB-2 as shown in panel A and mCh::TBG-1 in panel B, along with mCh::HIS-48 (histone 48), which appears more strongly in later blastomeres (Toya et al., 2010). A merge of A and B is shown in panel C, and a DIC image is shown in panel D. Strain TY3558 contains GFP::TBB-2 and GFP::H2B (histone 2B) as shown in panel E, with corresponding DIC image in panel F (this strain is not referenced). In all panels, anterior is to the left. Note the asymmetric distribution of one or both of these components at the two-cell stage as seen by a brighter cytoplasm in AB relative to P1 (panel E) and that interphase blastomeres in older embryos show a cortical tubulin network (asterisks in panels A and C). Both strains are available at the Caenorhabditis Genetics Center (CGC). Scale bar is 10 μm.
Evidence for both redundancy and specificity among C. elegans tubulins has come from studies of the tubulins that function in somatic tissues in processes such as neuronal polarity, synaptogenesis, and plasticity (Baran et al., 2010; Ou and Shen, 2011; Driscoll et al., 1989; Kurup et al., 2015). The broadly expressed tubulins, perhaps with some contribution from the more specialized family members, likely function as the predominant microtubule tracks for the movement, positioning, and anchoring of intracellular membranous organelles such as yolk granules (McNally et al., 2010) and nuclei (Tapley and Starr, 2013; see Section 4.3.15 for description of NOCA-1). In a process involving the polarity protein PAR-3 microtubules and components of centrosomes relocalize during establishment of intestinal epithelial polarity (Feldman and Priess, 2012). Under control of the formin EXC-6 (excretory cell abnormal) microtubules help organize tubulogenesis (Shaye and Greenwald, 2015). In the absence of centrosomes, microtubules can polymerize from and help remodel epidermal junctions, where they function alongside the LET-502 (Rho kinase) pathway during embryonic elongation (Quintin et al., 2016; Wang et al. 2015). Microtubules also form the tracks for dynein-based movement of chromosomes during oocyte cell division (Muscat et al., 2015). During differentiation of reproductive muscles microtubules undergo regulated changes in their behavior that are controlled by regulatory proteins and/or compartmentation of soluble monomer (Lacroix et al., 2014; Lacroix et al., 2016).
Two α- (TBA-1 and TBA-2) and two β- (TBB-1 and TBB-2) tubulins show some degree of redundancy and are major components of the mitotic spindle microtubules and of the axonal microtubules in motor neurons. A third β-tubulin, BEN-1, functions redundantly in neurons.
TBA-1 is expressed in the germline and is involved in meiotic and mitotic spindle assembly in the early embryo. It is partially redundant with TBA-2 as RNAi depletion of both results in embryonic lethality while RNAi or deletion of either causes little effect on embryonic viability (Lu and Mains, 2005; Phillips et al., 2004; Honda et al. 2017). TBA-1 is also broadly expressed in excitatory motor neurons (Table 2, Section 6.2), where it functions with DLK-1 (dual leucine zipper kinase), a mitogen-activated protein kinase kinase kinase, in synaptogenesis and remodeling events (Kurup et al., 2015). Dominant or semi-dominant missense alleles, but not deletion alleles, cause defects in cell division events and neuronal function. These molecular and cellular defects underlie embryonic lethality (O'Rourke et al. 2011) and uncoordination, respectively (Baran et al., 2010). Suppressors of the neuronal phenotypes caused by the combination of a dominant allele of TBA-1 and loss of DLK-1 include a neomorphic allele of TBB-2 and novel missense alleles of kinesin, dynein/dynactin, and the casein kinase TTBK-3 (tau tubulin kinase) (Kurup et al. 2017).
The primary structure of TBA-2 is very closely related to that of TBA-1 (98.5% identical, 99.5% similar), which likely provides the basis for redundancy. Sequence similarity extends through their CTTs, which are also the most similar to α-CTTs of other organisms. TBA-2 is expressed in the germline and in certain neurons (Table 2, Section 6.2), overlapping with TBA-1 in the VB and DB motor neurons and the PLM sensory neurons (Fukushige et al., 1993). Like TBA-1, TBA-2 functions in the mitotic spindle of the early embryo as dominant missense alleles cause embryonic lethality (Lu and Mains, 2005; Phillips et al., 2004). Both TBA-1 and TBA-2 are also expressed in touch receptor neurons, but they do not seem to be required for the structure or function of those cells (Lockhead et al. 2016). However, minor sequence differences between TBA-1 and TBA-2, which are mostly at the amino-terminus, result in differential sensitivity to the actions of the microtubule severing protein, katanin (MEI-1/MEI-2) and in differences in microtubule dynamics in the early embryo (Lu and Mains, 2005; Honda et al. 2017).
Like the broadly expressed α-tubulins, TBB-1 and TBB-2 (97.6% identical and 98.9% similar) both function in a partially redundant way in the mitotic spindle in embryonic blastomeres, and they interact differently with MEI-1/MEI-2 katanin (Lu et al., 2004; Wright and Hunter, 2003). Similarly, they are partially redundant in motor neurons as loss of either causes mild axonal outgrowth and synaptic defects similar to the dominant mutation of tba-1 (loss of both is lethal). However, loss of either enhances the defects caused by a dominant allele of tba-1 (Baran et al., 2010). Both are expressed in many unidentified neurons and other tissues, but rarely does loss of function of one cause any significant defect (Lockhead et al. 2016).
TBB-2 functions during microtubule-dependent processes in the early embryo such as spindle placement and rotation, where dominant missense alleles cause stronger defects than do null alleles (Ellis et al., 2004; Wright and Hunter, 2003, O'Rourke et al., 2011). Loss of TBB-2 causes an up-regulation of TBB-1 in the early embryo, providing a genetic mechanism for redundancy (Ellis et al., 2004). Quantitative expression analysis in the early embryo indicates that TBB-2 is more prevalent than TBB-1, and substitution experiments suggest that the particular tubulins present and their concentration contribute to diverse microtubule behaviors (Honda et al. 2017).
Large-scale RNAi screens indicate that TBB-2 is also required for P-granule localization (Updike and Strome, 2009). Along with TBA-2, TBB-2 is required for correct sexual specification of the male gonad (Kalis et al., 2010). It is broadly expressed in many somatic tissues and the germline (Hunt-Newbury et al., 2007; Lu et al., 2004; McKay et al., 2003), and regulated by the formin EXC-6, it also plays a role in excretory cell morphogenesis (Shaye and Greenwald, 2015).
Benomyl is an anti-mitotic benzimidazole compound that acts to inhibit dynamic instability of microtubules (Gupta et al., 2004; Singh et al., 2008). When wild-type C. elegans are reared in its presence, growth is slowed, movement is uncoordinated, and fewer neuronal processes emanate from ventral cord motor neurons. The ben-1 locus was identified in a screen for mutations that offer resistance to benomyl (Driscoll et al., 1989). Deletion of ben-1 and certain missense mutations confer resistance while causing no other gross abnormalities. Mutations that cause either resistance or hypersensitivity to benomyl have been linked to a β-tubulin in both nematodes and fungi (Nakaune and Nakano, 2007). ben-1 is broadly expressed in the nervous system (Figure 5), which supports the idea that it is involved in the neuronal control of locomotion. However, no cytological phenotypes have been reported. The structure of the CTT of BEN-1 is similar to TBB-4 (Figure 3), although it lacks a recognizable axonemal motif (see Section 2.2.4).
|
Figure 5. ben-1 is expressed in multiple neurons. A transcriptional fusion (after Boulin et al., 2006: PMID 18050449) of the ben-1 promoter to yellow fluorescent protein (YFP, from pPD136.64/L4817 in the Fire Lab vector kit 1999) reveals broad expression in sensory, motor, and likely interneurons. Scale bars = 100 μm in panels A and D and 10 μm in B, C, E, and F. Boxes in D indicate views shown in E and F. Strain to be available at the CGC.
C. elegans sense their environment with an array of chemo- and mechanosensory neurons such as the anterior amphid neurons and posterior phasmid neurons. The non-motile cilia at the tips of the dendrites in these neurons are built on a 9+0 axoneme structure, and they have become a model in which to study homologs of the ciliopathy genes in humans (see WormBook chapter The sensory cilia of Caenorhabditis elegans;)
TBA-5 is an axonemal α-tubulin expressed in amphid and phasmid sensory neurons where it is localized to cilia. It was isolated in a screen to identify mutants in ciliary dye-filling. A missense allele causes a Dyf (dye-filling defective) phenotype and shortened cilia while loss-of-function (deletion) mutations do not show defects in dye-filling (Hao et al., 2011). TBA-5, TBB-4, and perhaps additional axonemal tubulins are transported by intraflagellar transport (IFT) mechanisms involving a number of microtubule motor proteins (Hao et al., 2011). No behavioral defects have been associated with the perturbation of tba-5.
TBA-6 was identified as a ciliary tubulin by combining serial analysis of gene expression (SAGE) and microarray datasets. It is co-expressed with PKD-2 (a polycystin) in ciliated, male-specific sensory neurons (CEMs and RnBs) in addition to inner labial neurons found in both sexes. Mutant analysis indicated that it is required, along with TBB-4, for the normal structure of CEM cilia and the normal localization of PKD-2 to the distal tips of RnB cilia (Hurd et al., 2010). TBA-6 is also required for multiple aspects of the specialized structure and function of male-specific CEM neurons including axoneme ultrastructure, motor protein velocity, and extracellular vesicle function (Silva et al., 2017). The combination of these abnormalities likely underlies the male mating behavior defects shown by mutants (Hurd et al., 2010). TBA-6 mRNA is presumably expressed in the germline as it is found in the one-cell embryo. It is also expressed in other somatic tissues (Baleanu-Gogonea and Siddiqui, 2000), but deletion mutants are fertile and without obvious morphological abnormalities. Structurally, TBA-6 lacks a recognizable α-tubulin axonemal CTT found in other ciliary α-tubulins.
TBA-9 was also identified as a candidate ciliary α-tubulin by combining SAGE and microarray datasets (Hurd et al., 2010). It is expressed in many ciliated sensory neurons in the head of both sexes, often with TBB-4 (Table 2, Section 6.2). It is required for many normal behaviors including male mating, retraction after nose touch, locomotory rate/posture, exploratory behavior, and the basal slowing response upon contact with food. Deletion causes only subtle defects in the localization of signaling proteins (D. D. Hurd, unpublished results), although it is required for the normal distribution of TBB-4. Structurally, it lacks a recognizable axonemal CTT found in other ciliary α-tubulins (Hurd et al., 2010).
TBB-4 is a broadly expressed ciliary β-tubulin orthologous to ciliary β-tubulins in other species (Hao et al., 2011). It has a recognizable axonemal motif in its CTT, and it is expressed in many, but not all ciliated sensory neurons, including certain amphid, certain labial, and both classes of male tail ray neurons (Table 2, Section 6.2) where it is required for the normal localization of signaling proteins, such as PKD-2. TBB-4 is required for normal male mating and nose-touch behaviors (Hurd et al., 2010). It is enriched in the ciliary axoneme, and is transported by intraflagellar transport. Some missense mutations are more severe than null (deletion) mutations (Hao et al., 2011).
The MEC-12 α-tubulin and the MEC-7 β-tubulin were originally discovered in screens for mutants that did not respond to mechanical stimulation along the body (Chalfie and Sulston, 1981). They are both expressed in the six body wall touch receptor neurons (TRNs) sometimes referred to as the microtubule cells (ALML/R, PLML/R, AVM, and PVM). These sensory neurons contain cross-linked 15-protofilament microtubules that terminate near the plasma membrane (Chalfie and Thomson, 1982). MEC-12 is the only C. elegans α-tubulin with a lysine as aa40, a position that is frequently modified via post-translational acetylation (see Section 4.2.1). Antibodies to acetylated tubulin strongly label the touch receptor cells (Akella et al., 2010; Shida et al., 2010; Siddiqui et al., 1989).
MEC-12 and MEC-7 are required for the 15 protofilament microtubules in the TRNs, for mechanoreceptor currents, and for touch sensitivity (Bounoutas et al., 2009; O'Hagan et al., 2005). Mutations often cause the appearance of 11 protofilament microtubules, which can alter the distribution and/or expression of the components of the mechanosensation signal transduction pathway (Bounoutas et al., 2011; Bounoutas et al., 2009; Huang et al., 1995). Mutations in these two tubulins that cause the most severe defects are not null, but rather dominant or semi-dominant missense (antimorphic or neomorphic) alleles (Hsu et al., 2014; Savage et al., 1994; Kirszenblat et al., 2013). Loss of PTL-1 (see Section 4.3.1) causes an enhancement of touch insensitivity in double mutant combinations with certain alleles of mec-12 or mec-7 (Gordon et al., 2008).
TBA-7 is also expressed in the TRNs, but is not strictly required for their behavioral function nor their characteristic 15 protofilament microtubules. Instead it plays a role in microtubule dynamics in these cells as loss of function increases stability, causes ectopic neurite growth and resistance to cochicine in a manner similar to KLP-7 (Zheng et al., 2017).
TBA-4 has not been studied in detail. Data from RNAi experiments have suggested that TBA-4 may play a role in embryonic development or larval growth (Simmer et al., 2003). Its anatomic expression resembles that of tba-7 (see Table 2, Section 6.2, and Figure 6).
|
Figure 6. tba-7 is primarily expressed in the intestine. A transcriptional fusion of the tba-7 promoter to YFP reveals expression predominantly in the gut (see Table 2). It is also expressed in the pore cell of the excretory system (arrow in panel C, which is a high magnification view of the junction of the posterior pharynx and anterior intestine). Scale bars = 100 μm in panel A and 10 μm in B and C. Box in panel B indicates view in panel C. Strain to be available at the CGC.
The primary structure of TBA-8 is notable for its diverged CTT that has a phenylalanine as the last amino acid (Figure 3). Nearly all other α-tubulins have a tyrosine that is often removed after translation. Transcriptional reporters indicate that it is expressed in seam cells and unidentified neurons (D. D. Hurd and D. S. Portman, unpublished results). No loss of function analysis has been undertaken.
TBB-6 is the most structurally diverged β-tubulin as it lacks most of what is considered to be a CTT (typically acidic amino acids, see Figure 3). It appears to have arisen recently in the Caenhorhabditis lineage (Saunders et al. 2013). Its anatomic expression closely resembles that of tba-7 (Table 2, Section 6.2). Neither a deletion mutation nor RNAi depletion causes an overt phenotype (D. D. Hurd, unpublished results).
γ-tubulin functions in the nucleation and anchoring of microtubules. Antibody staining and translational GFP fusions show that γ-tubulin is a component of centrosomes (Bobinnec et al., 2000; Strome et al., 2001). It is also found at the base of cilia in amphid and phasmid sensory neurons and the apical cortex in the intestinal epithelium (Bobinnec et al., 2000). RNAi depletion of γ-tubulin causes polyploid germline cells and early blastomeres that contain a disorganized, ineffective mitotic spindle (Bobinnec et al., 2000; Strome et al., 2001). Too much γ-tubulin at the centrosome is caused by loss of the RNA-binding protein ATX-2 (ataxin-2), and this also brings about multiple defects in cell division (Stubenvoll et al., 2016). γ-tubulin is not absolutely required for microtubule nucleation, but instead is required for proper placement of microtubule minus ends within centrosomes and normal centriole structure (O'Toole et al., 2012).
The C. elegans genome does not contain orthologs of other tubulins (δ-, ϵ- or η-tubulin), which are found in some single-celled eukaryotes and vertebrates. These tubulins are perhaps involved in basal body and centriole function (Dutcher, 2003; Oakley, 2000).
Microtubules underlie a variety of essential cellular processes and are regulated by diverse mechanisms at many levels including biogenesis, heterodimer formation, nucleation, polymerization, depolymerization, post-translational modification, and association with other proteins (see Table 3, Section 6.3).
Newly synthesized tubulin (and actin) monomers pass through a series of chaperone complexes prior to being incorporated into dimers and eventually microtubules. The first of these is the prefoldin complex, which accepts nascent tubulin monomers from the ribosome and delivers them to the CCT (chaperonin-containing TCP-1) complex. The CCT complex finalizes folding of monomers and presents them to the tubulin folding cofactors A-E for dimerization. Tubulin degradation is thought to be mediated by the ubiquitin-proteasome pathway and other regulators/sensors of tubulin homeostasis, such as parkin or cofactor E-like proteins (Lundin et al., 2010).
PFD-1 through PFD-6 are subunits of the prefoldin complex, and CCT-1 through CCT-8 compose the CCT complex. Both transgenic fusion proteins and antibody staining indicate that they are expressed in nearly all cells with the most intense expression (of transgene) in body wall muscle, pharyngeal muscle, vulval muscle, hypodermal cells, and the somatic gonad. Inhibition by RNAi in the early embryo indicates that prefoldin, CCT, and TXDC-9 function are required for multiple events in the early embryo that are dependent on microtubule function in addition to distal tip cell migration (Leroux and Candido, 1997; Lundin et al., 2008; Ogawa et al., 2004; Srayko et al., 2005)). Post-embryonic RNAi depletion of cct-5 has shown that it is required for proper levels of tubulin and normal organization of intestinal actin and microvilli (Saegusa et al., 2014). The tubulin folding cofactors have not been studied in detail. Parkin has been implicated in the ubiquitination of tubulin in other systems, but the role for the broadly expressed C. elegans ortholog (PRD-1) in tubulin homeostasis has not been investigated (Springer et al., 2005).
Eukaryotic tubulins can be covalently modified after translation in a number of ways. These include tubulin-specific modifications such as removal/replacement of a terminal tyrosine (α-tubulin), addition of an acetyl group to an amino-terminal lysine (α-), addition of glycines or glutamates to residues in the CTT (both α- and β-). Tubulins also undergo more general modifications such as phosphorylation/dephosphorylation (β-) and palmitylation (α-). The most intensely studied modifications are those that are unique to tubulins (Hammond et al., 2008; Janke and Bulinski, 2011).
MEC-12 is the only α-tubulin with a lysine at position 40, suggesting that it is the only acetylated α-tubulin in C. elegans. Supporting this idea, antibodies that recognize acetylated tubulin intensely stain the body wall TRNs, and neurons implicated in nose mechanosensation that express MEC-12 (Shida et al., 2010; Siddiqui et al., 1989).
Acetylation is accomplished by two α-tubulin acetyltransferases (aTATs), MEC-17 and ATAT-2. They are expressed in TRNs and are required for acetylation of tubulin on the lumenal side of the microtubule wall in these cells. Acetylated α-tubulin is required for optimal touch sensation (Akella et al., 2010; Shida et al., 2010) and the structure and mechanical properties of the specialized 15 protofilament microtubules found in touch receptor neurons, although MEC-17 might play an additional role in microtubule structure (Cueva et al., 2012; Topalidou et al., 2012, Davenport et al., 2014).
Tubulins are glutamylated by tubulin tyrosine ligating enzymes (TTLLs). Some members of this family initiate while some elongate chains of glutamate. These chains are added to a glutamate residue in the primary structure of tubulin CTTs. TTLL-4 glutamylates microtubules in the amphid channel cilia and certain labial cilia (See Table 3, Section 6.3). Individual TTLL mutations cause little overt phenotypic effect, but a triple mutant of TTLL-4, TTLL-5, and TTLL-11 eliminates glutamylation and causes reduced male mating efficiency (Chawla et al. 2016).
Certain members of the cytosolic carboxypeptidase (CCPP) family remove glutamate chains from tubulin. Both CCPP-1 and CCPP-6 have been shown to suppress poly- or hyper-glutamylation (Kimura et al., 2010; O'Hagan et al., 2011). Proper levels of glutamylation are required for axonemal microtubule ultrastructure, normal velocity of motor proteins, localization of signaling proteins, and certain behaviors that require input from ciliated neurons (O'Hagan et al., 2011).
Microtubules are found in their numerous cellular contexts in either highly dynamic and transient structures, such as the spindle during mitosis, more stable structures, such as the axoneme supporting a cilia, or in variably stable structures, such as neuronal microtubules during development or after injury. These higher order structures and/or changes in dynamic behavior are regulated by a collection of microtubule-associated proteins that can promote growth/stabilize, promote shrinkage/destabilize, sever, bundle, bind to plus or minus ends, or perform some combination of these functions.
tau and other similar MAPs bind to and stabilize microtubules. In vertebrates, dysfunction of tau causes disassociation from microtubules and self-aggregation, which underlies a broad class of neuronal pathologies known collectively as tauopathies (Sabbagh and Dickey, 2016). C. elegans has a single identified protein that is similar to mammalian tau (Goedert et al., 1996; McDermott et al., 1996). PTL-1 (protein with tau-like repeats) is most highly conserved in both structure and function to tau and other MAPs in the carboxyl-terminal microtubule binding repeats (Hashi et al., 2016; Gordon et al., 2008). It is most prominently expressed in mechanoreceptor cells. Loss of PTL-1 causes no overt phenotypic effect, but ptl-1 mutations dominantly enhance the loss of touch sensation caused by lack of either MEC-7 or MEC-12 (Goedert et al., 1996; Gordon et al., 2008). Loss of both PTL-1 and spectrin (UNC-70) also causes a rearrangement of microtubule bundles, which sensitizes TRN neurites to mechanical damage (Krieg et al. 2017). PTL-1 is required for the normal behavior of microtubule motor proteins and the distribution of synaptic components in touch receptor cells (Tien et al., 2010).
EMAP (echinoderm microtubule-associated protein) family members bind to and can either stabilize (Houtman et al., 2007) or destabilize microtubules (Eichenmüller et al., 2002). Consensus domains across family members include ~6 WD40 repeats and another domain specific to the EMAP-like proteins, the ELP domain. C. elegans has a single member of the family, ELP-6 (Suprenant et al., 2000), which has a predicted amino terminal coiled domain found in some of the vertebrate orthologs (Hueston et al., 2008). It has been shown to bind to microtubules and is expressed in a number of cell types including body wall muscle, sensory neurons (IL1, male ray neurons, TRNs), vulval muscles, the spermathecal valve cell, intestinal cells, and seam cells (Hueston et al., 2008). Loss of ELP-1 through mutation or RNAi causes defects in touch reception, and in combination with loss of the dystrophin homolog DYS-1, causes paralysis and death (Hueston and Suprenant, 2009).
The transforming acidic coiled-coil (TACC-1) family of proteins functions in the regulation of microtubule length by promoting growth (Mortuza et al, 2014). The C. elegans protein TAC-1 consists solely of the TACC-1 domain, while orthologous proteins have more complex and larger structures (Bellanger and Gönczy, 2003; Le Bot et al., 2003). TAC-1 accumulates in the centrosomes of dividing blastomeres (dependent upon ZYG-9) where it physically associates and functions with ZYG-9 and ZYG-8 to regulate microtubule length, pronuclear migration, and spindle positioning as a polymerizing factor (Bellanger et al., 2007; Bellanger and Gönczy, 2003; Srayko et al., 2003).
Doublecortin kinases bind to microtubules to stabilize and/or straighten them (Jean et al., 2012). ZYG-8 is the ortholog of the human protein DCLK (doublecortin-like kinase), as it contains a doublecortin domain and a kinase domain similar to calcium/calmodulin-regulated kinases. ZYG-8 localizes to microtubules in the mitotic spindle in early blastomeres (and in cultured mammalian cells), where it functions to promote microtubule growth required for proper spindle placement (Gönczy et al., 2001). It is also expressed in certain neurons where it is required for process outgrowth, locomotion, and touch sensitivity (Bellanger et al., 2012). ZYG-8 physically associates with TAC-1 through its doublecortin domain (Bellanger et al., 2007) and with EFA-6 during regeneration after axonal injury (see Section 4.3.8).
The XMAP215 (Xenopus microtubule-associated protein) family of microtubule binding proteins is composed of three TOG (tumor overexpressed gene) domains that interact with tubulin dimers and promote assembly (Al-Bassam and Chang, 2011). ZYG-9 is the C. elegans member, originally discovered in screens for mutations that alter polarity in the early embryo (Kemphues et al., 1986). It was subsequently shown to be required for spindle function (Matthews et al., 1998). ZYG-9 localizes to the centrosome in dividing blastomeres (dependent upon TAC-1), where it functions with ZYG-8 and TAC-1 to regulate the length of microtubules (Bellanger et al., 2007; Bellanger and Gönczy, 2003; Srayko et al., 2003).
The CLASP (cytoplasmic linker associated protein) family of microtubule regulators also contains TOG domains. They function in microtubule stabilization by promoting rescue of shrinking microtubules (Al-Bassam and Chang, 2011). CLS-1, CLS-2, and CLS-3 are the members of the CLASP family in C. elegans. RNAi depletion causes defects in nuclear rotation, maintenance of spindle length, and spindle placement in the one-cell embryo, likely due to fewer astral microtubules. CLS-2 is found on the mitotic spindle, centrosomes, and kinetochores in dividing cells in early embryogenesis and in the cytoplasm of interphase cells (Espiritu et al, 2012).
Crescerins (crescere: to grow) are another family of TOG domain proteins that bind to and regulate microtubules in cilia. This family is represented by CHE-12 in C. elegans, and is part of a larger family that includes the CLASPs (CLS-1, -2 and -3) and ch-TOG/XMAP215 (ZYG-9) proteins. CHE-12 localizes to the cilia of the amphid channel neurons and the phasmid neurons. Loss of function through mutation causes altered cilia structure, inability to take up lipophilic dyes, and failure in chemotaxis (Bacaj et al., 2008; Das et al., 2015).
Members of the EFA-6 (exchange factor for ARF6 family GTPases) protein family function to limit microtubule growth. These proteins are composed of a Sec7 guanine nucleotide exchange factor domain, a pleckstrin homology (PH) domain, and regions of coiled-coil. C. elegans EFA-6 is found in the cortex of blastomeres where loss of function through mutation or RNAi treatment causes an increase in the total number of microtubules, an increase in the fraction of the population that is growing, and an increase in microtubule length (Chen et al., 2011; O'Rourke et al., 2010). EFA-6 is also found dynamically rearranging from a smooth distribution to punctae of microtubule minus ends during axotomy-induced regeneration in PLMs. This is controlled by an 18 amino acid N-terminal motif through which it also makes physical association with TAC-1 and ZYG-8. Loss-of-function mutations cause neurite overgrowth after injury (Chen et al., 2015).
RMD-1 through -6 is a family of proteins that might function in the regulation of microtubule dynamics. They all contain predicted coiled-coil domains and have orthologs in vertebrates, although little is known about the vertebrate homologs. Only RMD-1 has been studied in detail; it appears to function in the attachment of microtubules to kinetochores as depletion through RNAi causes delayed anaphase and incorrectly segregated DNA during mitosis. RMD-1 localizes to the spindle of the one-cell embryo and binds to microtubules in vitro (Oishi et al., 2007).
Katanins are a family of microtubule severing proteins (Roll-Mecak and McNally, 2010). MEI-1 is the p60 catalytic subunit (AAA ATPase) of C. elegans katanin, and MEI-2 is the p80 activator/regulator of MEI-1. They were originally identified in screens for defects in meiosis and mitosis (Clark-Maguire and Mains, 1994; Mains et al., 1990). They are maternally expressed and dependently colocalize in the meiotic spindle of oocytes (Srayko et al., 2000). It was originally thought that microtubule severing activity was required for functional meiotic spindles (Srayko et al., 2006), but recent analysis of specific point mutations suggests that MEI-1 might also bind/bundle microtubules (McNally and McNally, 2011; Connolly et al., 2014; McNally et al., 2014). Levels of MEI-1 decrease after meiosis due to the activity of the MEL-26 E3 ubiquitin ligase (Johnson et al., 2009; Pintard et al., 2003).
Like katanins, spastins are microtubule severing proteins in the AAA ATPase family of enzymes (Roll-Mecak and McNally, 2010). SPAS-1 was identified via homology in a search for proteins related to spastin, the human protein that is defective in a form of spastic paraplegia. SPAS-1 is reportedly expressed in a broad variety of tissues in worms. A precise in vivo function is not evident from studies of a deletion mutation, which causes pleitropic defects in growth, vulval development, and reproduction (Matsushita-Ishiodori et al., 2007). Biochemically, SPAS-1 forms hexamers, and its ATPase activity is stimulated by microtubules. Conserved basic amino acids interact with acidic CTTs of tubulin, and a conserved tryptophan in SPAS-1 is required for microtubule severing (Matsushita-Ishiodori et al., 2009).
KLP-7 is a kinesin-like protein of the MCAK (mitotic centromere-associated kinesin) family, which functions to depolymerize microtubules in many systems (Sanhaji et al., 2011). It is found at the kinetochore in the blastomeres of the early embryo (Oegema et al., 2001). KLP-7 is phosphorylated by Aurora kinases, and it regulates nucleation rate; loss of klp-7 expression through RNAi or mutation increases the number of astral and midzone microtubules while decreasing growth rate (Srayko et al., 2005; Han et al., 2015). KLP-7 also functions to establish a bipolar meiotic spindle in the absence of centrosomes (Connolly et al., 2015). In adult touch receptor neurons, KLP-7 functions to maintain a dynamic population of microtubules in normal cells and to inhibit microtubule upregulation in axotomized cells (Ghosh-Roy et al. 2012).
Microtubule end-binding proteins (also known as tip-interacting proteins or +TIPs) recognize and bind to a structural feature at the plus ends of microtubules (Jiang and Akhmanova, 2011). EBP-1 and EBP-2 fusion proteins (driven by the pie-1 promoter) decorate the growing plus ends of microtubules in the spindle of early embryos (Motegi et al., 2006; Srayko et al., 2005). EBP-1 is required for efficient axon regrowth in PLM neurons after laser axotomy (Chen et al., 2011). Perturbation by mutation or RNAi does not cause severe phenotypes, perhaps due to redundancy, and analysis of anatomic expression from native promoters for these three paralogs has not been reported.
Members of the CAMSAP (calmodulin-regulated spectrin-associated protein)/Nezha/patronin family decorate and stabilize minus ends of microtubules (Akhmanova and Hoogenraad, 2015). The proteins share a conserved structure, which includes a calponin homology domain, a coiled-coil domain(s), the namesake CAMSAP (also called CKK) domain, and a carboxyl-terminal microtubule binding domain. PTRN-1 in C. elegans is broadly expressed in neurons and has been shown to be associated with microtubules in puncta in nerve and muscle cells (Marcette et al. 2014; Richardson et al., 2014; Chuang et al. 2014). It is not required in general for normal movement, sensation, or reproduction. The effects of loss-of-function mutations, such as overgrowth or ectopic neurite formation, are more readily observed in microtubule-sensitized situations such as exposure to the depolymerizing drug colchicine, or in specific developmental contexts such as neuronal remodeling in touch receptor neurons or after injury. Loss of DLK-1 suppresses the subtle defects caused by loss of PTRN-1, suggesting they may work in the same pathway (Marcette et al. 2014; Richardson et al., 2014; Chuang et al. 2014).
Ninein proteins function to anchor and perhaps stabilize and/or nucleate microtubules through association with the minus end (Srivatsa et al. 2015). The C. elegans homolog of vertebrate ninein-like proteins is NOCA-1, encoded by a complex locus that produces several variants. It was originally identified as a gene that phenocopied γ-tubulin defects. A deletion causes abnormal positioning of germline nuclei and associated sterility, with no other obvious phenotypes. NOCA-1 works with γ-tubulin and PTRN-1 to establish non-centrosomal microtubule arrays that position nuclei and/or provide structural integrity (along with LET-502) to the hypodermis (Quintin et al., 2016). NOCA-1 and γ-tubulin colocalize, and NOCA-1 and PTRN-1 bind to microtubules in extracts (Wang et al. 2015).
Cylicin was originally discovered in the cytoskeletal calyx that surrounds part of the nucleus in the head of mammalian sperm; structurally it is characterized by numerous small repeats of the charged amino acids KKD/E (Hess et al. 1993). C. elegans has two highly conserved proteins, CYLC-1 and CYLC-2, that are related to mammalian cylicin and potentially to stathmin. Inhibition by RNAi causes altered tubulin concentrations and misregulated growth of microtubules in a region of uterine muscle cells, suggesting that these proteins function in the intracellular compartmentation of tubulin levels (Lacroix et al., 2016). No anatomic expression pattern has yet been reported.
As a genetic and cytological model in which to study a family of closely related structural proteins, C. elegans provides evidence for both redundancy and specificity in the use of α- and β-tubulins. In support of redundancy, mutagenesis screening has produced a collection of missense alleles that change conserved amino acids in tubulin (e.g., in the tba-1, tba-5, tbb-2, mec-12, mec-7, and tbb-4 genes) that often cause more severe defects than deletions/nulls. This means that it is often more deleterious to incorporate an altered tubulin into a microtubule than it is to incorporate a wild-type version of another tubulin. In addition, genetic and/or RNAi elimination of multiple mitotic tubulins (tba-1, tba-2, tbb-1, and tbb-2) is required to disrupt embryogenesis, further supporting redundancy among tubulins. Nevertheless, specific expression patterns (Figure 7), the presence of variant CTTs, and the quantifiable cytological and behavioral defects caused by the loss of single tubulins (e.g., tba-5, tba-6, tba-9, mec-12, tbb-4, or mec-7) all support the idea that tubulins can be specialized for certain developmental and physiological contexts. Whether tubulins in C. elegans are regulated in response to environmental change has not been explored.
|
Figure 7. Summary of the somatic expression of the α- and β-tubulins. Venn diagrams depict the anatomic expression of the tubulin family members in non-germline tissues/organs. The type of data used to place a particular tubulin in a particular area is depicted using color. See text for references.
C. elegans provides the ability to understand tubulin biology from the molecular to the organismal level. Worm tubulins will continue to be discovered playing roles in diverse and unforeseen biological processes, whether through de novo mutagenesis screening, systems level analysis, or novel approaches. At the molecular level, new alleles can continue to shed insight into the structure and function of the tubulin dimer and the myriad other proteins that interact with tubulin, either soluble or assembled. Both forward and reverse genetic screens and searching for modifiers of existing phenotypes, especially among paralogs expressed in the same tissues and/or with regulatory factors, allows understanding of tubulin usage and regulation within an organism. Technological advances (e.g., studying transport velocities of cellular components in neurons or other tissues), a strength of the nematode model system, should provide a higher resolution and more precise quantitative description of the role of tubulin in metazoan biology.
| Gene | Allele | Protein change | Strain | Mutation class | Reference |
|---|---|---|---|---|---|
| tba-1 | or346 | S377F | EU1135 | Dominant | Philips et al., 2004; O'Rourke et al., 2011 |
| tba-1 | or594 | S377F | EU1161 | Semi-dominant | O'Rourke et al., 2011 |
| tba-1 | ju89 | G414R | Dominant | Baran et al., 2010 | |
| tba-2 | sb116 | E22K | Intragenic revertant of sb51 | Lu, C. et al., 2005 | |
| tba-2 | sb117 | E69K | Intragenic revertant of sb51 | Lu, C. et al., 2005 | |
| tba-2 | sb51 | S168Y | HR505 | Dominant | Lu, C. et al., 2005 |
| tba-2 | sb27 | E194K | HR973 | Dominant | Lu, C. et al., 2005 |
| tba-2 | sb25 | E277K | HR899 | Dominant | Lu, C. et al., 2005 |
| mec-12 | gk170196 | P32S | Recessive, Neomorph | Zheng et al., 2017 | |
| mec-12 | u76 | D69N | Recessive, Antimorph | Fukushige et al., 1999; Zheng et al., 2017 | |
| mec-12 | u1016 | E97K | Recessive, Antimorph | Zheng et al., 2017 | |
| mec-12 | u950, gk672907 | S140F | Recessive, Antimorph | Zheng et al., 2017 | |
| mec-12 | gk600523 | G142E | Recessive | Zheng et al., 2017 | |
| mec-12 | u1021, e1607 | G144S | Semi-dominant (haplo-insufficient), Recessive, Antimorph | Hsu et al., 2014; Bountonas et al. 2011; Bountonas et al., 2009; Zheng et al., 2017 | |
| mec-12 | u94 | G148S | Semi-dominant | Gu et al., 1996; Bounoutas et al., 2009 | |
| mec-12 | u67 | M154I | Temperature-sensitive dominant | Gordon et al., 2008; Bounoutas et al., 2009 | |
| mec-12 | u204 | S178F | Chalfie and Au 1989; Bounoutas et al., 2009 | ||
| mec-12 | u279 | T179I | Gu et al., 1996 | ||
| mec-12 | u50, e1605 | H192Y | CB3284 | Recessive, Loss of function | Bountonas et al., 2009; Bountonas et al., 2011 |
| mec-12 | gk915672 | E196K | Recessive, Neomorph | Zheng et al., 2017 | |
| mec-12 | u1041 | G246E | Recessive, Loss of function | Zheng et al., 2017 | |
| mec-12 | u917 | V260I | Recessive, Neomorph | Zheng et al., 2017 | |
| mec-12 | u159 | C295Y | Semi-dominant | Gu et al., 1996; Bounoutas et al., 2009 | |
| mec-12 | gk515972 | V323I | Recessive, Neomorph | Zheng et al., 2017 | |
| mec-12 | u241, u1019 | G354E | Dominant, Gain of function, Recessive, Antimorph | Fukushige et al, 1999; Bountonas et al., 2011; Bountonas et al., 2009; Zheng et al., 2017 | |
| mec-12 | u63 | E415K | Recessive, Loss of function | Fukushige et al., 1999; Bounoutas et al., 2011; Zheng et al., 2017 | |
| mec-12 | gm379 | G416E | Semi-dominant, Recessive, Loss of function | Hsu et al., 2014 | |
| tba-5 | qj14 | A19V | Gain-of-Function | Hao et al., 2011 | |
| tbb-2 | or600 | G140E | EU1588 | Semi-dominant | O'Rourke et al., 2011 |
| tbb-2 | or362 | G141E | EU858 | Semi-dominant, Dominant | Ellis et al, 2004; Lu et al., 2004 |
| tbb-2 | qt1 | E198K | HC48 | Gain-of-Function | Wright et al., 2003 |
| tbb-2 | t1623 | V313M | GE2255 | Semi-dominant | Wright et al., 2003 |
| tbb-2 | sb26 | E439K | HR1060 | Semi-dominant, Gain of function | Lu, et.al., 2004; Shaye et al., 2015 |
| mec-7 | e1506 | M1I | CB3270 | Recessive | Savage et al., 1994; Zheng et al., 2017 |
| mec-7 | u319 | S25F | Semi-dominant, Antimorph | Savage et al., 1994; Zheng et al., 2017 | |
| mec-7 | u305, u1020 | G34S | Recessive, Loss of function | Savage et al., 1994; Sze et al., 1997; Zheng et al., 2017 | |
| mec-7 | u58, u223 | P61L | Semi-dominant, Antimorph | Savage et al., 1994 | |
| mec-7 | u249 | P61S | Semi-dominant, Antimorph | Savage et al., 1994; Zheng et al., 2017 | |
| mec-7 | u430 | A97V | Recessive, Antimorph | Savage et al., 1994; Zheng et al., 2017 | |
| mec-7 | u9, u440 | W101stop | Recessive | Savage et al., 1994; Zheng et al., 2017 | |
| mec-7 | u222 | G109E | Semi-dominant, Antimorph | Savage et al., 1994; Zheng et al., 2017 | |
| mec-7 | u429, u433 | G141E | Recessive, Loss of function | Savage et al., 1994; Zheng et al., 2017 | |
| mec-7 | gk595364 | G141R | Recessive, Loss of function | Zheng et al., 2017 | |
| mec-7 | u275 | G148R | Recessive, Loss of function | Savage et al., 1994; Zheng et al., 2017 | |
| mec-7 | gk895768 | T149I | Recessive, Neomorph | Zheng et al., 2017 | |
| mec-7 | gk286003 | R162Q | Recessive, Neomorph | Zheng et al., 2017 | |
| mec-7 | e1343, u127, u957 | P171L | CB1477; CB2217 | Semi-dominant, Antimorph | Savage et al., 1994; Zheng et al., 2017: PMID |
| mec-7 | u911 | P171S | Dominant, Antimorph | Zheng et al., 2017 | |
| mec-7 | u1056 | S172P | Recessive, Loss of function | Zheng et al., 2017 | |
| mec-7 | u48 | S176F | Semi-dominant, Antimorph | Savage et al., 1994; Zheng et al., 2017 | |
| mec-7 | u449 | V179A | Semi-dominant, Antimorph | Savage et al., 1994; Zheng et al., 2017 | |
| mec-7 | u10 | S188F | Recessive, Loss of function | Savage et al., 1994; Zheng et al., 2017 | |
| mec-7 | u225 | T214P | Recessive, Loss of function | Savage et al., 1994; Zheng et al., 2017 | |
| mec-7 | ky852 | P220S | Kirszenblat et.al. 2013 | ||
| mec-7 | u262 | N226Y | Semi-dominant, Antimorph | Savage et al., 1994; Zheng et al., 2017 | |
| mec-7 | gk286002 | P234S | Recessive, Loss of function | Zheng et al., 2017 | |
| mec-7 | u283 | P243L | Dominant, Antimorph | Savage et al., 1994; Bountonas et al., 2011: 21368137; Zheng et al., 2017 | |
| mec-7 | u129, u958 | G244S | Dominant, Antimorph | Savage et al., 1994; Zheng et al., 2017 | |
| mec-7 | n434 | N247I | TU300 | Dominant, Antimorph | Savage et al., 1994; Zheng et al., 2017 |
| mec-7 | u162 | D249N | Dominant, Antimorph | Savage et al., 1994; Zheng et al., 2017 | |
| mec-7 | u427 | V255M | Semi-dominant | Savage et al., 1994 | |
| mec-7 | e1505 | G269D | Recessive, Loss of function | Savage et al., 1994; Zheng et al., 2017 | |
| mec-7 | u142 | Q279stop | Recessive | O'Hagan et.al 2005; Zheng et al., 2017 | |
| mec-7 | u178, u431 | Q280stop | Recessive | Sze et al., 1997 | |
| mec-7 | e1527 | V286D | CB3276 | Dominant | Savage et al., 1994; Zheng et al., 2017 |
| mec-7 | u451 | Q292P | Semi-dominant, Antimorph | Savage et al., 1994 | |
| mec-7 | u445 | M300V | Semi-dominant, Antimorph | Savage et al., 1994; Zheng et al., 2017 | |
| mec-7 | u98 | M300T | Semi-dominant, Antimorph | Savage et al., 1994; Zheng et al., 2017 | |
| mec-7 | u1058 | A302T | Semi-dominant, Neomorph | Zheng et al., 2017 | |
| mec-7 | u278 | C303Y | Semi-dominant, Neomorph | Savage et al., 1994; Zheng et al., 2017 | |
| mec-7 | gk286000 | A314V | Semi-dominant, Antimorph | Zheng et al., 2017 | |
| mec-7 | e1522 | F317I | Recessive, Loss of function | Savage et al., 1994; Zheng et al., 2017 | |
| mec-7 | u234 | R318Q | Recessive, Loss of function | Savage et al., 1994; Zheng et al., 2017 | |
| mec-7 | u88 | R318stop | Recessive | Sze et al., 1997 | |
| mec-7 | u955 | A352T | Dominant, Antimorph | Zheng et al., 2017 | |
| mec-7 | u910, gk373602 | P357L | Dominant, Antimorph | Zheng et al., 2017 | |
| mec-7 | u956 | P358L | Dominant, Antimorph | Zheng et al., 2017 | |
| mec-7 | u143 | G369R | Semi-dominant | Savage et al., 1994 | |
| mec-7 | u428 | G369E | Recessive, Loss of function | Gu et al., 1996; Zheng et al., 2017 | |
| mec-7 | u1017 | L377F | Recessive, Neomorph | Zheng et al., 2017 | |
| mec-7 | u1059 | R380S | Recessive, Neomorph | Zheng et al., 2017 | |
| mec-7 | u18 | A393T | Dominant, Antimorph | Savage et al., 1994; Zheng et al., 2017 | |
| mec-7 | gk285997 | A393V | Dominant, Antimorph | Zheng et al., 2017 | |
| mec-7 | u173 | G402R | Recessive | Savage et al., 1994 | |
| mec-7 | u170 | E407L | Recessive, Neomorph | Savage et al., 1994; Zheng et al., 2017 | |
| mec-7 | u1060 | E410K | Semi-dominant, Antimorph | Zheng et al., 2017 | |
| mec-7 | u382 | E410:Tc5 | Recessive | Savage et al., 1994 | |
| mec-7 | u388 | E410:Tc5 | Recessive | Savage et al., 1994 | |
| mec-7 | u443 | Recessive | Savage et al., 1994 | ||
| mec-7 | u448 | Recessive | Savage et al., 1994 | ||
| mec-7 | u453 | Recessive | Savage et al., 1994 | ||
| mec-7 | u80 | Recessive | Savage et al., 1994 | ||
| mec-7 | u136 | Semi-dominant | Savage et al., 1994 | ||
| tbb-4 | sa127 | L253F | SP1742 | Gain of function | Hao et al., 2011 |
| ben-1 | e1880 | CB3474 | Semi-dominant | Driscoll et al., 1989 | |
| ben-1 | e1910 | Semi-dominant | Driscoll et al., 1989 | ||
| ben-1 | e1911 | Semi-dominant | Driscoll et al., 1989 | ||
| ben-1 | u102 | Semi-dominant | Driscoll et al., 1989 | ||
| ben-1 | u116 | Semi-dominant | Driscoll et al., 1989 | ||
| ben-1 | u134 | Semi-dominant | Driscoll et al., 1989 | ||
| ben-1 | u462 | Semi-dominant | Driscoll et al., 1989 | ||
| ben-1 | u463 | Semi-dominant | Driscoll et al., 1989 | ||
| tbb-6 | oxTi207 | EG7952 | |||
| tbg-1 | t1465 | A401V | GE2959 | Hannack et al., 2002 |
| Locus | Gene Model | Expression | Cellular, developmental, behavioral roles | Selected references |
|---|---|---|---|---|
| tba-1 | F26E4.8 | ALM, PLM, AVM, PVM, VA/VB/DA/DB motor neurons, multiple other neurons, germline, early embryo | cellular architecture, meiosis/mitosis, neuronal pathfinding, synaptogenesis | Lu and Mains, 2005; Phillips et al., 2004; Baran et al., 2010 |
| tba-2 | C47B2.3 | pharynx, intestine, VB/DB motor neurons, PLM, ALA, other anterior neurons, germline, early embryo | cellular architecture, meiosis/mitosis | Lu and Mains, 2005; Phillips et al., 2004 |
| mec-12 | C44B11.3 | ALM, PLM, AVM, PVM, other neurons | sensory neuron dendritic microtubules, mechanosensation | Chalfie and Sulston, 1981; Bounoutas et al., 2009; O'Hagan et al., 2005; Bounoutas et al., 2011 |
| tba-4 | F44F4.11 | intestine and other tissues | unknown/redundant | D. D. Hurd, unpublished (expression) |
| tba-5 | F16D3.1 | PHA, PHB, amphid neurons, perhaps other sensory neurons | sensory neuron axonemal microtubules, chemosensation and/or mechanosensation | Hao et al., 2011 |
| tba-6 | F32H2.9 | IL2, CEM, HSN, RnB, HOB | sensory neuron axonemal/dendritic microtubules, chemosensation and/or mechanosensation | Hurd et al., 2010 |
| tba-7 | T28D6.2 | ALM, PLM, AVM, PVM, intestine, and excretory pore cell (Figure 5) | microtubule dynamics | Figure 6 and Zheng et al., 2017 |
| tba-8 | ZK899.4 | seam cells, neurons | unknown/redundant | D. D. Hurd and D. S. Portman, unpublished (expression) |
| tba-9 | F40F4.5 | CEP, OLQ, ASI, ADF, AFD, AWA, AWC, ASE, ADE, URX, RIG, PDE, PQR, RnA, HOA, various motor neurons, other neurons, and neuronal support cells | axonal/axonemal/dendritic microtubules, chemosensation and/or mechanosensation | Hurd et al., 2010 |
| tbb-1 | K01G5.7 | germline, early embryo | cellular architecture, meiosis/mitosis | Lu et al., 2004; Wright and Hunter, 2003 |
| tbb-2 | C36E8.5 | germline, early embryo, neurons, excretory cell | cellular architecture, meiosis/mitosis, neuronal pathfinding, synaptogenesis, | SLu et al., 2004; Wright and Hunter, 2003 |
| mec-7 | ZK154.3 | ALM, PLM, AVM, PVM, FLP, PVD, BDU | sensory neuron dendritic microtubules, mechanosensation | Chalfie and Sulston, 1981; Bounoutas et al., 2009; O'Hagan et al., 2005; Bounoutas et al., 2011 |
| tbb-4 | B0272.1 | CEP, OLQ, CEM, ASK, ADL, ASI, ASJ, ASH, AWB, AWA, AWC, ASE, ASG, FLP, HSN, PDE, PQR, PHA, PHB, RnA, RnB, HOA, vulval muscles | axonemal/dendritic microtubules, chemosensation and/or mechanosensation | Hurd et al., 2010; Hao et al., 2010 |
| ben-1 | C54C6.2 | motor, sensory, and interneurons | axonal pathfinding, sensitivity to benzimidazole | Figure 5 and Driscoll et al., 1989 |
| tbb-6 | T04H1.9 | intestine and other tissues | unknown/redundant | D. D. Hurd, unpublished (expression) |
| tbg-1 | F58A4.8 | early embryo, sensory neurons, other tissues | microtubule organization/nucleation, spindle organization, nuclei positioning, tissue integrity, embryonic elongation | Bobinnec et al., 2000; Strome et al., 2001; O'Toole et al., 2012; Quintin et al., 2016; Wang et al. 2015 |
| Locus | Sequence | Expression | Molecular, cellular, developmental, and/or behavioral roles | Selected references |
|---|---|---|---|---|
| pfd-1, pfd-2, pfd-3, pfd-4, pfd-5, pfd-6 | C08F8.1, H20J04.5, T06G6.9, B0035.4, R151.9, F21C3.5 | ubiquitous | protein folding, a-tubulin levels, microtubule dynamics, cell division, cell migration | Lundin et al., 2008; |
| cct-1, cct-2, cct-3, cct-4, cct-5, cct-6, cct-7, cct-8 | T05C12.7, T21B10.7, F54A3.3, K01C8.10, C07G2.3, F01F1.8, T10B5.5, Y55F3AR.3 | ubiquitous | protein folding, a-tubulin levels, cell division, cell migration, actin localization, formation/maintenance of microvilli in intestine | Saegusa et al., 2014 |
| mec-17, atat-2 | F57H12.7, W06B11.1 | both: ALM, AVM, PLM, and PVM atat-2 only: OLQ, CEP, PVR | acetylation of a-tubulin, microtubule architecture and stability, mechanosensation | Cueva et al., 2012; Topalidou et al., 2012, Davenport et al., 2014 |
| ttll-4, ttll-5, ttll-9, ttll-11, ttll-12, ttll-15 | ZK1128.6, C55A6.2, F25C8.5, H23L24.3, D2013.9, K07C5.7 | amphid and other sensory neurons, body wall muscles, motor neurons | glutamylation of tubulins | Kimura et al., 2010; O'Hagan et al., 2011 |
| ccpp-1, ccpp-6 | F56H1.5, EEED8.6 | ASE, ASG, ASH, ASI, ASK, ASL, ADF, ADL, AWA, AWB, AWC, AFD, IL2, CEM, RnB1-RnB9, HOB | deglutamylation of tubulins, microtubule architecture and stability, motor protein activity, cilia stability | Kimura et al., 2010; O'Hagan et al., 2011 |
| ptl-1 | F42G9.9 | ALM, AVM, PLM, PVM (early larva only), other neurons | motor protein activity, egg hatching, mechanosensation | Goedert et al., 1996; Gordon et al., 2008; Tien et al., 2010 |
| elp-1 | F38A6.2 | ALM, AVM, PLM, PVM, IL1, male tail ray neurons, body wall muscle, vulval muscles, spermathecal valve cell, intestine, and seam cells | associates with microtubules; mechanosensation, motililty and egg-laying (along with DYS-1) | Hueston et al., 2008; Hueston and Suprenant, 2009 |
| tac-1 | Y54E2A.3 | centrosomes of blastomeres | associates with ZYG-8 and ZYG-9, polymerizes/stabilizes microtubules, pronuclear migration, spindle positioning | Bellanger and Gönczy, 2003; Srayko et al., 2003 |
| zyg-8 | Y79H2A.11 | centrosomes of blastomeres, ALM, AVM, PLM, PVM, VD and DD motor neurons | associates with TAC-1 and ZYG-9, polymerizes/stabilizes microtubules, pronuclear migration, spindle positioning, cell polarity, locomotion, mechanosensation | Gönczy et al., 2001; ; Bellanger et al., 2012 |
| zyg-9 | F22B5.7 | centrosomes of blastomeres | associates with TAC-1 and ZYG-8, polymerizes/stabilizes microtubules, pronuclear migration, spindle positioning | Matthews et al., 1998; Bellanger et al., 2007 |
| cls-1, cls-2, cls-3 | C07H6.3, R107.6, ZC84.3 | centrosomes, kinetochores, and spindle of blastomeres | stabilize microtubules, spindle length and positioning | Espiritu et al, 2012 |
| che-12 | B0024.8 | ASE, ASG, ASH, ASI, ASJ, ASK, PHA, PHB | stabilizes axonal microtubules, cilia structure, chemosensation | Bacai et al., 2008; Das et al., 2015 |
| efa-6 | Y55D9A.1 | germline, blastomeres, neurons | associates with TAC-1 and ZYG-8, inhibits axon regrowth after injury | O'Rourke et al., 2010; Chen et al., 2011; Chen et al., 2015 |
| rmd-1, rmd-2, rmd-3, rmd-4, rmd-5, rmd-6 | T05G5.7, C27H6.4, B0491.3, F36H12.11, T23B3.3, R13H9.1 | germline, blastomeres | associates with microtubules, associates with AIR-2 (aurora B kinase), kinetochore-microtubule attachment | Oishi et al., 2007 |
| mei-1, mei-2 | T01G9.5, F57B10.12 | germline, blastomeres | severs and/or bundles microtubules, meiotic spindle formation and maintenance | McNally and McNally, 2011; Connolly et al., 2014; McNally et al., 2014 |
| spas-1 | C24B5.2 | ubiquitous | severs microtubules, reproduction | Matsushita-Ishiodori et al., 2009 |
| klp-7 | K11D9.1 | kinetochores, blastomeres, neurons | depolymerizes microtubules, meiotic spindle assembly | Srayko et al., 2005; Ghosh-Roy et al. 2012; Connolly et al., 2015 |
| ebp-1, ebp-2, ebp-3 | Y59A8B.7, VW02B12L.3, Y59A8B.9 | unknown | interact with plus ends of microtubules, axon regrowth after injury | Motegi et al., 2006; Chen et al., 2011 |
| ptrn-1 | F35B3.5 | neurons | polymerizes microtubules, axon remodeling and regrowth after injury | Marcette et al. 2014: ; Richardson et al., 2014: ; Chuang et al. 2014: |
| noca-1 | T09E8.1 | hypodermis, neurons, intestine, pharynx | polymerizes (non centrosomal) microtubules, tissue integrity (with γ-tubulin and PTRN-1), nuclear positioning | Wang et al. 2015; Quintin et al., 2016 |
I thank Douglas Portman (University of Rochester) for initiating my interest in the C. elegans tubulins and the continued use of equipment in his laboratory. I also thank Noveera Ahmed (St. John Fisher College) for critically reading versions of the manuscript, Nicholas Hoffend and Katelyn Giardino (St. John Fisher College undergraduates) for making the DNA constructs that led to Figure 5 and Figure 6, and Michelle Price (St. John Fisher College Lavery Library) for the reference collecting and Textpresso searching that led to Table 1 (Section 6.1). Finally, I acknowledge the community of C. elegans researchers for the production of seminal and important data in the field of tubulin biology.
Akella, J.S., Wloga, D., Kim, J., Starostina, N.G., Lyons-Abbott, S., Morrissette, N.S., Dougan, S.T., Kipreos, E.T., and Gaertig, J. (2010). MEC-17 is an α-tubulin acetyltransferase. Nature 467, 218-222. Abstract Article
Akhmanova, A., and Hoogenraad, C.C. (2015). Microtubule minus-end-targeting proteins. Curr. Biol. 25, R162-R171. Abstract Article
Al-Bassam, J., and Chang, F. (2011). Regulation of microtubule dynamics by TOG-domain proteins XMAP215/Dis1 and CLASP. Trends Cell Biol. 21, 604-614. Abstract Article
Alushin, G.M., Lander, G.C., Kellogg, E.H., Zhang, R. Baker, D. and Nogales, E. (2014). High resolution microtubule structures reveal the structural transitions in αβ-tubulin upon GTP hydrolysis. Cell 157, 1117-1129. Abstract Article
Bacaj, T., Lu, Y., and Shaham, S. (2008). The conserved proteins CHE-12 and DYF-11 are required for sensory cilium function in Caenorhabditis elegans. Genetics 178, 989-1002. Abstract Article
Baleanu-Gogonea, C. and Siddiqui, S.S. (2000). Molecular cloning and three-dimensional structure prediction of a novel α-tubulin in Caenorhabditis elegans. J. Cell. Mol. Med. 4, 183-195. Abstract Article
Bajaj, M. and Srayko, M. (2013). Laulimalide induces dose-dependent modulation of microtubule behaviour in the C. elegans embryo. PLoS One 8, e71889. Abstract Article
Baran, R., Castelblanco, L., Tang, G., Shapiro, I., Goncharov, A., and Jin, Y. (2010). Motor neuron synapse and axon defects in a C. elegans α-tubulin mutant. PLoS One 5, e9655. Abstract Article
Bellanger, J.M., Carter, J. C., Phillips, J.B., Canard, C., Bowerman, B., and Gönczy, P. (2007). ZYG-9, TAC-1 and ZYG-8 together ensure correct microtubule function throughout the cell cycle of C. elegans embryos. J. Cell Sci. 120, 2963-2973. Abstract Article
Bellanger, J.M., Cueva, J.G., Baran, R., Tang, G., Goodman, M.B., and Debant, A. (2012). The doublecortin-related gene zyg-8 is a microtubule organizer in Caenorhabditis elegans neurons. J. Cell. Sci. 125, 5417-5427. Abstract Article
Bellanger, J.M., and Gönczy, P. (2003). TAC-1 and ZYG-9 form a complex that promotes microtubule assembly in C. elegans embryos. Curr. Biol. 13, 1488-1498. Abstract Article
Bienkowska, D. and Cowan, C R. (2012). Centrosomes can initiate a polarity axis from any position within one-cell C. elegans embryos. Curr. Biol. 22, 583-589. Abstract Article
Bobinnec, Y., Fukuda, M., and Nishida, E. (2000). Identification and characterization of Caenorhabditis elegans γ-tubulin in dividing cells and differentiated tissues. J. Cell Sci. 113, 3747-3759. Abstract Article
Bounoutas, A., Kratz, J., Emtage, L., Ma, C., Nguyen, K.C., and Chalfie, M. (2011). Microtubule depolymerization in Caenorhabditis elegans touch receptor neurons reduces gene expression through a p38 MAPK pathway. Proc. Natl. Acad. Sci. U. S. A. 108, 3982-3987. Abstract Article
Bounoutas, A., O'Hagan, R., and Chalfie, M. (2009). The multipurpose 15-protofilament microtubules in C. elegans have specific roles in mechanosensation. Curr. Biol. 19, 1362-1367. Abstract Article
Chalfie, M., and Sulston, J. (1981). Developmental genetics of the mechanosensory neurons of Caenorhabditis elegans. Dev. Biol. 82, 358-370. Abstract Article
Chalfie, M., and Thomson, J.N. (1982). Structural and functional diversity in the neuronal microtubules of Caenorhabditis elegans. J. Cell Biol. 93, 15-23. Abstract Article
Chalfie, M., and Au, M. (1989). Genetic control of differentiation of the Caenorhabditis elegans touch receptor neurons. Science 243, 1027-1033. Abstract Article
Chawla, D.G., Shah, R.V., Barth, Z.K., Lee, J.D., Badecker, K.E., Naik, A., Brewster, M.M., Salmon, T.P., and Peel, N. (2016). Caenorhabditis elegans glutamylating enzymes function redundantly in male mating. Biol. Open 5, 1290-1298. Abstract Article
Cheerambathur, D.K., Gassman, R., Cook, B., Oegema, K., and Desai, A. (2013). Crosstalk between microtubule attachment complexes ensures accurate chromosome segregation. Science 342, 1239-1242 Abstract Article
Chen, L., Wang, Z., Ghosh-Roy, A., Hubert, T., Yan, D., O'Rourke, S., Bowerman, B., Wu, Z., Jin, Y., and Chisholm, A.D. (2011). Axon regeneration pathways identified by systematic genetic screening in C. elegans. Neuron 71, 1043-1057. Abstract Article
Chen, L., Chuang, M., Koorman, T., Boxem, M., Jin, Y., and Chisholm, A.D. (2015). Axon injury triggers EFA-6 mediated destabilization of axonal microtubules via TACC and doublecortin like kinase. eLife 4, doi: 10.7554/eLife.08695. Abstract Article
Chuang, M., Goncharov, A., Wang, S., Oegema, K., Jin Y., and Chisholm, A.D. (2014). The microtubule minus-end-binding protein patronin/PTRN-1 is required for axon regeneration in C. elegans. Cell Rep. 9, 874-883. Abstract Article
Clark-Maguire, S., and Mains, P.E. (1994). mei-1, a gene required for meiotic spindle formation in Caenorhabditis elegans, is a member of a family of ATPases. Genetics 136, 533-546. Abstract Article
Cleveland, D.W. (1987). The multitubulin hypothesis revisited: what have we learned? J. Cell Biol. 104, 381-383. Abstract Article
Connolly, A.A., Osterberg, V., Christensen, S., Price, M., Lu, C., Chicas-Cruz, K., Lockery, S., Mains, P.E., Bowerman, B. (2014). Caenorhabditis elegans oocyte meiotic spindle pole assembly requires microtubule severing and the calponin homology domain protein ASPM-1. Mol. Biol. Cell 25, 1298-1311. Abstract Article
Connolly, A.A., Sugioka, K., Chuang, C., Lowry, J., Bowerman, B. KLP-7 acts through the Ndc80 complex to limit pole number in C. elegans oocyte meiotic spindle assembly. (2015). J. Cell Biol. 210, 917-932. Abstract Article
Cowan, C.R. and Hyman, A.A. (2004). Centrosomes direct cell polarity independently of microtubule assembly in C. elegans embryos. Nature 431, 92-96. Abstract Article
Cueva, J.G., Hsin, J., Huang, K.C., and Goodman, M.B. (2012). Postranslational acetylation of α-tubulin constrains protofilament number in native microtubules. Curr. Biol. 22, 1066-1074. Abstract Article
Das, A., Dickinson, D.J., Wood, C.C., Goldstein, B., and Slep, K.C. (2015). Crescerin uses a TOG domain array to regulate microtubules in the primary cilium. Mol. Biol. Cell 26, 4248-4264. Abstract Article
Davenport, A.M., Collins, L.N., Chiu, H. Minor, P.J. Sternberg, P.M., and Hoelz, A. (2014). Structural and functional characterization of the α-tubulin acetyltransferase MEC-17. J. Mol. Biol. 426, 2605-2616. Abstract Article
Driscoll, M., Dean, E., Reilly, E., Bergholz, E., and Chalfie, M. (1989). Genetic and molecular analysis of a Caenorhabditis elegans β-tubulin that conveys benzimidazole sensitivity. J. Cell Biol. 109, 2993-3003. Abstract Article
Duerr, J. (2013). Antibody staining in C. elegans using “freeze-cracking”. J. Vis. Exp. 80, doi: 10.3791/50664. Abstract Article
Dubey, J., Ratnakaran, N., Koushika, S.P. (2015). Neurodegeneration and microtubule dynamics: death by a thousand cuts. Front. Cell. Neurosci. 9, 343. Abstract Article
Dutcher, S.K. (2003). Long-lost relatives reappear: identification of new members of the tubulin superfamily. Curr. Opin. Microbiol. 6, 634-640. Abstract Article
Eichenmüller, B., Everley, P., Palange, J., Lepley, D., and Suprenant, K.A. (2002). The human EMAP-like protein-70 (ELP70) is a microtubule destabilizer that localizes to the mitotic apparatus. J. Biol Chem. 277, 1301-1309. Abstract Article
Ellis, G.C., Phillips, J.B., O'Rourke, S., Lyczak, R., and Bowerman, B. (2004). Maternally expressed and partially redundant β-tubulins in Caenorhabditis elegans are autoregulated. J. Cell. Sci. 117, 457-464. Abstract Article
Erickson, H.P. (1998). Atomic structures of tubulin and FtsZ. Trends Cell Biol. 8, 133-137. Abstract Article
Espiritu, E.B., Krueger, L.E., Ye, A. and Rose, L.S. (2012). CLASPs function redundantly to regulate astral microtubules in the C. elegans embryo. Dev. Biol. 368, 242-254. Abstract Article
Feldman, J.L., and Priess, J.R. (2012). A role for the centrosome and PAR-3 in the hand-off of microtubule organizing center function during epithelial polarization. Curr. Biol. 22, 572-582. Abstract Article
Fukushige, T., Siddiqui, Z.K., Chou, M., Culotti, J.G., Gogonea, C.B., Siddiqui, S.S., and Hamelin, M. (1999). MEC-12, an α-tubulin required for touch sensitivity in C. elegans. J. Cell Sci. 112, 395-403. Abstract Article
Fukushige, T., Yasuda, H., and Siddiqui, S.S. (1993). Molecular cloning and developmental expression of the alpha-2 tubulin gene of Caenorhabditis elegans. J. Mol. Biol. 234, 1290-1300. Abstract Article
Fukushige, T., Yasuda, H., and Siddiqui, S.S. (1995). Selective expression of the tba-1 α tubulin gene in a set of mechanosensory and motor neurons during the development of Caenorhabditis elegans. Biochim. Biophys. Acta 1261, 401-416. Abstract Article
Goedert, M., Baur, C.P., Ahringer, J., Jakes, R., Hasegawa, M., Spillantini, M.G., Smith, M.J., and Hill, F. (1996). PTL-1, a microtubule-associated protein with tau-like repeats from the nematode Caenorhabditis elegans. J. Cell Sci. 109, 2661-2672. Abstract Article
Gogonea, C.B., Gogonea, V., Ali, Y. M., Merz, K.M., Jr., and Siddiqui, S.S. (1999). Computational prediction of the three-dimensional structures for the Caenorhabditis elegans tubulin family. J. Mol. Graph. Model. 17, 90-100. Abstract Article
Gönczy, P., Bellanger, J.-M., Kirkham, M., Pozniakowski, A., Baumer, K., Phillips, J.B., and Hyman, A.A. (2001). zyg-8, a gene required for spindle positioning in C. elegans, encodes a doublecortin-related kinase that promotes microtubule assembly. Dev. Cell 1, 363-375. Abstract Article
Gönczy, P. and Rose, L.S. (2005). Asymmetric cell division and axis formation in the embryo . WormBook, ed. The C. elegans Research Community, WormBook, doi: 10.1895/wormbook.1.30.1, http://www.wormbook.org. Article
Gordon, P., Hingula, L., Krasny, M.L., Swienckowski, J.L., Pokrywka, N.J., and Raley-Susman, K. M. (2008). The invertebrate microtubule-associated protein PTL-1 functions in mechanosensation and development in Caenorhabditis elegans. Dev. Genes Evol. 218, 541-551. Abstract Article
Ghosh-Roy, A., Goncharov, A., Jin, Y., Chisholm, A.D. (2012). Kinesin-13 and tubulin posttranslational modifications regulate microtubule growth in axon regeneration. Dev. Cell, 23, 716-728. Abstract Article
Gu, G., Caldwell, G.A., and Chalfie, M. (1996). Genetic interactions affecting touch sensitivity in Caenorhabiditis elegans. Proc. Natl. Acad. Sci. U. S. A. 93, 6577-6582. Abstract Article
Gupta, K., Bishop, J., Peck, A., Brown, J., Wilson, L., and Panda, D. (2004). Antimitotic antifungal compound benomyl inhibits brain microtubule polymerization and dynamics and cancer cell proliferation at mitosis, by binding to a novel site in tubulin. Biochemistry 43, 6645-6655. Abstract Article
Hammond, J.W., Cai, D., and Verhey, K.J. (2008). Tubulin modifications and their cellular functions. Curr. Opin. Cell Biol. 20, 71-76. Abstract Article
Han, X., Adames, K., Sykes, E.M., and Srayko, M. (2015). The KLP-7 residue S546 is a putative Aurora kinase site required for microtubule regulation at the centrosome in C. elegans. PLoS One 10, e0132593. Abstract Article
Hannack, A., Oegema, K., Kirckham, M., Gönczy, P., Habermann, B., and Hyman, A.A. (2002). The kinetically dominant assembly pathway for centrosomal asters in Caenorhabditis elegans is γ-tubulin dependent. J. Cell Biol. 157, 591-602. Abstract Article
Hao, L., Thein, M., Brust-Mascher, I., Civelekoglu-Scholey, G., Lu, Y., Acar, S., Prevo, B., Shaham, S., and Scholey, J.M. (2011). Intraflagellar transport delivers tubulin isotypes to sensory cilium middle and distal segments. Nat. Cell Biol. 13, 790-798. Abstract Article
Hashi, Y., Kotani, S., and Adachi, T. (2016). A nematode microtubule-associated protein, PTL-1, closely resembles its mammalian counterparts in overall molecular architecture. Biosci. Biotech. Biochem. 80, 1107-1113. Abstract Article
Hayashi, H., Kimura, K., and Kimura, A. (2012). Localized accumulation of tubulin during semi-open mitosis in the Caenorhabditis elegans embryo. Mol. Biol. Cell 23, 1688-1699. Abstract Article
Hess, H., Heid, H., and Franke, W.W. (1993). Molecular characterization of mammalian cylicin, a basic protein of the sperm head cytoskeleton. J. Cell Biol. 122, 1043-1052. Abstract Article
Honda, Y., Tsuchiya, K., Sumiyoshi, E., Haruta, N., and Sugimoto, A. (2017). Tubulin isotype substitution revealed that isotype combination modulates microtubule dynamics in C. elegans embryos. J. Cell Sci. 130, 1652-1661. Abstract Article
Houtman, S.H., Rutteman, M., De Zeeuw, C.I., and French, P.J. (2007). Echinoderm microtubule-associated protein like protein 4, a member of the echinoderm microtubule-associated protein family, stabilizes microtubules. Neuroscience 144, 1373-1382. Abstract Article
Hsu, J.M., Chen, C.H., Chen, Y.C., McDonald, K.L., Gurling, M., Lee, A., Garriga, G., and Pan, C.L. (2014). Genetic analysis of a novel tubulin mutation that redirects synaptic vesicle targeting and causes neurite degeneration in C. elegans. PLoS Genet. 10, e1004715. Abstract Article
Huang, M., Gu, G., Ferguson, E.L., and Chalfie, M. (1995). A stomatin-like protein necessary for mechanosensation in C. elegans. Nature 378, 292-295. Abstract Article
Hueston, J.L., Herren, G.P., Cueva, J.G., Buechner, M., Lundquist, E.A., Goodman, M.B., and Suprenant, K.A. (2008). The C. elegans EMAP-like protein, ELP-1 is required for touch sensation and associates with microtubules and adhesion complexes. BMC Dev. Biol. 8, 110. Abstract Article
Hueston, J.L., and Suprenant, K.A. (2009). Loss of dystrophin and the microtubule-binding protein ELP-1 causes progressive paralysis and death of adult C. elegans. Dev. Dyn. 238, 1878-1886. Abstract Article
Hunt-Newbury, R., Viveiros, R., Johnsen, R., Mah, A., Anastas, D., Fang, L., Halfnight, E., Lee, D., Lin, J., Lorch, A., et al. (2007). High-throughput in vivo analysis of gene expression in Caenorhabditis elegans. PLoS Biol. 5, e237. Abstract Article
Hurd, D.D., Miller, R.M., Nunez, L., and Portman, D.S. (2010). Specific α- and β-tubulin isotypes optimize the functions of sensory cilia in Caenorhabditis elegans. Genetics 185, 883-896. Abstract Article
Ingerson-Mahar M. and Gitai, Z. (2012). A growing family: the expanding universe of the bacterial cytoskeleton. FEMS Microbiol. Rev. 36, 256-266. Abstract Article
Inglis, P.N., Ou, G., Leroux, M.R., and Scholey, J.M. (2007). The sensory cilia of Caenorhabditis elegans. WormBook, ed. The C. elegans Research Community, WormBook, doi: 10.1895/wormbook.1.126.2, http://www.wormbook.org. Article
Janke, C., and Bulinski, J.C. (2011). Post-translational regulation of the microtubule cytoskeleton: mechanisms and functions. Nat. Rev. Mol. Cell Biol. 12, 773-786. Abstract Article
Jean, D.C., Baas, P.W., and Black, M.M. (2012). A novel role for doublecortin and doublecortin-like kinase in regulating growth cone microtubules. Hum. Mol. Genet. 21, 5511-5527 Abstract Article
Jiang, K., and Akhmanova, A. (2011). Microtubule tip-interacting proteins: a view from both ends. Curr. Opin. Cell Biol. 23, 94-101. Abstract Article
Johnson, J.L., Lu, C., Raharjo, E., McNally, K., McNally, F.J., and Mains, P.E. (2009). Levels of the ubiquitin ligase substrate adaptor MEL-26 are inversely correlated with MEI-1/katanin microtubule-severing activity during both meiosis and mitosis. Dev. Biol. 330, 349-357. Abstract Article
Kalis, A.K., Kroetz, M.B., Larson, K.M., and Zarkower, D. (2010). Functional genomic identification of genes required for male gonadal differentiation in Caenorhabditis elegans. Genetics 185, 523-535. Abstract Article
Kemphues, K J., Wolf, N., Wood, W.B., and Hirsh, D. (1986). Two loci required for cytoplasmic organization in early embryos of Caenorhabditis elegans. Dev. Biol. 113, 449-460. Abstract Article
Kimura, Y., Kurabe, N., Ikegami, K., Tsutsumi, K., Konishi, Y., Kaplan, O I., Kunitomo, H., Iino, Y., Blacque, O.E., and Setou, M. (2010). Identification of tubulin deglutamylase among Caenorhabditis elegans and mammalian cytosolic carboxypeptidases (CCPs). J. Biol. Chem. 285, 22936-22941. Abstract Article
Kirszenblat, L., Neumann, B., Coakley, S., and Hilliard, M.A. (2013). A dominant mutation in mec-7/β-tubulin affects axon development and regeneration in Caenorhabditis elegans neurons. Mol. Biol. Cell. 24, 285-296. Abstract Article
Krieg, M., Stühmer, J., Cueva, J.G., Fetter, R., Spilker, K., Cremers, D., Shen, K., Dunn, A.R., and Goodman, M.B. (2017). Genetic defects in β-spectrin and tau sensitize C. elegans axons to movement-induced damage via torque-tension coupling. eLife 6, e20172. Abstract Article
Kurup, N., Yan, D., Goncharov, A., and Jin, Y. (2015). Dynamic microtubules drive circuit rewiring in the absence of neurite remodeling. Curr. Biol. 25, 1594-1605. Abstract Article
Kurup, N., Yan, D., Kono, K., Jin, Y. (2017). Differential regulation of polarized synaptic vesicle trafficking and synapse stability in neural circuit rewiring in Caenorhabditis elegans. PLoS Genet. 13, e1006844. Abstract Article
Lacroix, B., Bourdages, K.G., Dorn, J.F., Ihara, S., Sherwood, D.R., Maddox, P.S., and Maddox, A.S. (2014). In situ imaging in C. elegans reveals developmental regulation of microtubule dynamics. Dev. Cell 29, 203-216. Abstract Article
Lacroix, B., Ryan, J., Dumont, J., Maddox, P.S., and Maddox, A.S. (2016). Identification of microtubule growth deceleration and its regulation by conserved and novel proteins. Mol. Biol. Cell 27, 1479-1487. Abstract Article
Le Bot, N., Tsai, M.C., Andrews, R.K., and Ahringer, J. (2003). TAC-1, a regulator of microtubule length in the C. elegans embryo. Curr. Biol. 13, 1499-1505. Abstract Article
Leroux, M.R., and Candido, E.P. (1997). Subunit characterization of the Caenorhabditis elegans chaperonin containing TCP-1 and expression pattern of the gene encoding CCT-1. Biochem. Biophys. Res. Commun. 241, 687-692. Abstract Article
Lockhead, D., Schwarz, E.M, O'Hagan, R., Bellotti, S., Krieg, M., Barr, M.M., Dunn, A.R., Sternberg, P.W., and Goodman, M.B. (2016). The tubulin repertoire of Caenorhabditis elegans sensory neurons and its context-dependent role in process outgrowth. Mol. Biol. Cell 27, 3717-3728. Abstract Article
Lu, C., and Mains, P.E. (2005). Mutations of a redundant α-tubulin gene affect Caenorhabditis elegans early embryonic cleavage via MEI-1/katanin-dependent and -independent pathways. Genetics 170, 115-126. Abstract Article
Lu, C., Srayko, M., and Mains, P E. (2004). The Caenorhabditis elegans microtubule-severing complex MEI-1/MEI-2 katanin interacts differently with two superficially redundant β-tubulin isotypes. Mol. Biol. Cell 15, 142-150. Abstract Article
Lundin, V.F., Leroux, M R., and Stirling, P.C. (2010). Quality control of cytoskeletal protein and human disease. Trends Biochem. Sci. 35, 288-297. Abstract Article
Lundin, V.F., Srayko, M., Hyman, A.A., and Leroux, M R. (2008). Efficient chaperone-mediated tubulin biogenesis is essential for cell division and cell migration in C. elegans. Dev. Biol. 313, 320-334. Abstract Article
Mains, P.E., Kemphues, K.J., Sprunger, S.A., Sulston, I.A., and Wood, W.B. (1990). Mutations affecting the meiotic and mitotic divisions of the early Caenorhabditis elegans embryo. Genetics 126, 593-605. Abstract Article
Marcette, J.D., Chen, J.J., and Nonet, M.L. (2014). The Caenorhabditis elegans microtubule minus-end binding homolog PTRN-1 stabilizes synapses and neurites. eLife 3, e01637. Abstract Article
Marston, D.J., and Goldstein, B. (2006). Symmetry breaking in C. elegans: another gift from the sperm. Dev. Cell 11, 273-274. Abstract Article
Matsushita-Ishiodori, Y., Yamanaka, K., Hashimoto, H., Esaki, M., and Ogura, T. (2009). Conserved aromatic and basic amino acid residues in the pore region of Caenorhabditis elegans spastin play critical roles in microtubule severing. Genes Cells 14, 925-940. Abstract Article
Matsushita-Ishiodori, Y., Yamanaka, K., and Ogura, T. (2007). The C. elegans homologue of the spastic paraplegia protein, spastin, disassembles microtubules. Biochem. Biophys. Res. Commun. 359, 157-162. Abstract Article
Matthews, L.R., Carter, P., Thierry-Mieg, D., and Kemphues, K. (1998). ZYG-9, a Caenorhabditis elegans protein required for microtubule organization and function, is a component of meiotic and mitotic spindle poles. J. Cell Biol. 141, 1159-1168. Abstract Article
McDermott, J.B., Aamodt, S., and Aamodt, E. (1996). ptl-1, a Caenorhabditis elegans gene whose products are homologous to the tau microtubule-associated proteins. Biochemistry 35, 9415-9423. Abstract Article
McKay, S.J., Johnsen, R., Khattra, J., Asano, J., Baillie, D.L., Chan, S., Dube, N., Fang, L., Goszczynski, B., Ha, E., et al. (2003). Gene expression profiling of cells, tissues, and developmental stages of the nematode C. elegans. Cold Spring Harbor Symp. Quant. Biol. 68, 159-169. Abstract Article
McNally, K.P., Martin, J.L., Ellefson, M. and McNally, F.J. (2010). Kinesin-dependent transport results in polarized migration of the nucleus in oocytes and inward movement of yolk granules in meiotic embryos. Dev. Biol. 339, 126-140. Abstract Article
McNally, K.P., and McNally, F.J. (2011). The spindle assembly function of Caenorhabditis elegans katanin does not require microtubule-severing activity. Mol. Biol. Cell. 22, 1550-1560. Abstract Article
McNally, K., Berg, E., Cortes, D.B., Hernandez, V., Mains, P.E., and McNally, F.J. (2014). Katanin maintains meiotic metaphase chromosome alignment and spindle structure in vivo and has multiple effects on microtubules in vitro. Mol. Biol. Cell 25, 1037-1049. Abstract Article
Mitchison, T., and Kirschner, M. (1984). Dynamic instability of microtubule growth. Nature 312, 237-242. Abstract Article
Mortuza, G.B., Cavazza, T., Garcia-Mayoral, M.F., Hermida, D., Peset, I., Pedrero, J.G., Merino, N., Blanco, F.J., Lyngsø, J., Bruix, M., et al. (2014). XTACC3-XMAP215 association reveals an asymmetric interaction promoting microtubule elongation. Nat. Commun. 5, 5072. Abstract Article
Motegi, F., Velarde, N.V., Piano, F., and Sugimoto, A. (2006). Two phases of astral microtubule activity during cytokinesis in C. elegans embryos. Dev. Cell. 10, 509-520. Abstract Article
Motegi, F., Zonies, S., Hao, Y., Cuenca, A.A., Griffin, E., and Seydoux, G. (2011). Microtubules induce self-organization of polarized PAR domains in C. elegans zygotes. Nat. Cell Biol. 13, 1361-1367. Abstract Article
Müller, R.U., Zank, S., Fabretti, F., and Benzing, T. (2011). Caenorhabditis elegans, a model organism for kidney research: from cilia to mechanosensation and longevity. Curr. Opin. Nephrol. Hypertens. 20, 400-408. Abstract Article
Müller-Reichert, T., Greenan, G., O'Toole, E., and Srayko, M. (2010). The elegans of spindle assembly. Cell. Mol. Life Sci. 67, 2195-2213. Abstract Article
Muscat, C.C., Torre-Santiago, K.M., Tran, M.V., Powers, J.A., and Wignall, S.M. (2015). Kinetochore-independent chromosome segregation driven by lateral microtubule bundles. eLife 4, e06462. Abstract Article
Nakaune, R., and Nakano, M. (2007). Benomyl resistance of Colletotrichum acutatum is caused by enhanced expression of β-tubulin 1 gene regulated by putative leucine zipper protein CaBEN1. Fungal Genet. Biol. 44, 1324-1335. Abstract Article
Nogales, E., Whittake, M., Milligan, R.A., and Downing, K.H. (1999). High-resolution model of the microtubule. Cell 96, 79-88. Abstract Article
Oegema, K., and Hyman, A.A. (2006). Cell division. WormBook, ed. The C. elegans Research Community, WormBook, doi: 10.1895/wormbook.1.72.1, http://www.wormbook.org. Article
Oegema, K., Desai, A., Rybina, S., Kirkham, M. and Hyman, A.A. (2001) Functional analysis of kinetochore assembly in Caenorhabditis elegans. J. Cell Biol. 153, 1209-1226. Abstract Article
Ogawa, S., Matsubayashi, Y., and Nishida, E. (2004). An evolutionarily conserved gene required for proper microtubule architecture in Caenorhabditis elegans. Genes Cells 9, 83-93. Abstract Article
O'Hagan, R., Chalfie, M., and Goodman, M.B. (2005). The MEC-4 DEG/ENaC channel of Caenorhabditis elegans touch receptor neurons transduces mechanical signals. Nat. Neurosci. 8, 43-50. Abstract Article
O'Hagan, R., Piasecki, B.P., Silva, M., Phirke, P., Nguyen, K.C., Hall, D.H., Swoboda, P., and Barr, M.M. (2011). The tubulin deglutamylase CCPP-1 regulates the function and stability of sensory cilia in C. elegans. Curr. Biol. 21, 1685-1694. Abstract Article
Oishi, K., Okano, H., and Sawa, H. (2007). RMD-1, a novel microtubule-associated protein, functions in chromosome segregation in Caenorhabditis elegans. J. Cell Biol. 179, 1149-1162. Abstract Article
O'Rourke, S.M., Christensen, S.N., and Bowerman, B. (2010). Caenorhabditis elegans EFA-6 limits microtubule growth at the cell cortex. Nat. Cell Biol. 12, 1235-1241. Abstract Article
O'Rourke, S.M., Carter, C., Carter, L., Christensen, S.N., Jones, M.P., Nash, B., Price, M.H., Turnbull, D.W., Garner, A.R., Hamill, D.R., et al. (2011). A survey of new temperature-sensitive, embryonic-lethal mutations in C. elegans: 24 alleles of thirteen genes. PLoS One 6, e16644. Abstract Article
O'Toole, E., Greenan, G., Lange, K.I., Srayko, M., and Müller-Reichert, T. (2012). The role of γ-tubulin in centrosomal microtubule organization. PLoS One 7, e29795. Abstract Article
Ou, C.Y., and Shen, K. (2011). Neuronal polarity in C. elegans. Dev. Neurobiol. 71, 554-566. Abstract Article
Phillips, J.B., Lyczak, R., Ellis, G.C., and Bowerman, B. (2004). Roles for two partially redundant α-tubulins during mitosis in early Caenorhabditis elegans embryos. Cell Motil. Cytoskel. 58, 112-126. Abstract Article
Pintard, L., Willis, J.H., Willems, A., Johnson, J.L., Srayko, M., Kurz, T., Glaser, S., Mains, P.E., Tyers, M., Bowerman, B., and Peter, M. (2003). The BTB protein MEL-26 is a substrate-specific adaptor of the CUL-3 ubiquitin-ligase. Nature 425, 311-316. Abstract Article
Popodi, E.M., Hoyle, H.D., Turner, F.R., Xu, K., Kruse, S., and Raff, E.C. (2008). Axoneme specialization embedded in a “generalist” β-tubulin. Cell Motil. Cytoskel. 65, 216-237. Abstract
Quintin, S., Wang, S., Pontabry, J., Bender, A., Robin, F., Hyenne, V., Landmann, F., Gally, C., Oegema, K., and Labouesse, M. (2016). Non-centrosomal epidermal microtubules act in parallel to LET-502/ROCK to promote C. elegans elongation. Development 143, 160-173. Abstract Article
Rice, L.M., Montabana, E.A. and Agard, D.A. (2008). The lattice as allosteric effector: structural studies of αβ- and γ-tubulin clarify the role of GTP in microtubule assembly. Proc. Natl. Acad. Sci. U. S. A. 105, 5378-5383. Abstract Article
Richardson, C.E., Spilker, K.A., Cueva, J.G., Perrino, J., Goodman, M.B., and Shen, K. (2014). PTRN-1, a microtubule minus end-binding CAMSAP homolog, promotes microtubule function in Caenorhabditis elegans neurons. eLife 3, e01498. Abstract Article
Roll-Mecak, A., and McNally, F.J. (2010). Microtubule-severing enzymes. Curr. Opin. Cell Biol. 22, 96-103. Abstract Article
Sabbagh, J.J., and Dickey, C.A. (2016) The metamorphic nature of the tau protein: dynamic flexibility comes at a cost. Front. Neurosci. 10, 3. Abstract Article
Saegusa, K., Sato, M., Sato, K., Nakajima-Shimada, J., Harada, A., and Sato, K. (2014). C. elegans chaperonin CCT/TRiC is required for actin and tubulin biogenesis and microvillus formation in intestinal epithelial cells. Mol. Biol. Cell. 25, 3095-3104. Abstract Article
Sanhaji, M., Friel, C.T., Wordeman, L., Louwen, F., and Yuan, J. (2011). Mitotic centromere-associated kinesin (MCAK): a potential cancer drug target. Oncotarget 2, 935-947. Abstract Article
Saunders, G.I., Wasmuth, J.D., Beech, R., Laing, R., Hunt, M., Naghra, H., Cotton, J.A., Berriman, M., Britton, C., and Gilleard, J.S. (2013). Characterization and comparative analysis of the complete Haemonchus contortus β-tubulin gene family and implications for benzimidazole resistance in strongylid nematodes. Int. J. Parasitol. 43, 465-475. Abstract Article
Savage, C., Xue, Y., Mitani, S., Hall, D., Zakhary, R., and Chalfie, M. (1994). Mutations in the Caenorhabditis elegans β-tubulin gene mec-7: effects on microtubule assembly and stability and on tubulin autoregulation. J. Cell Sci. 107, 2165-2175. Abstract Article
Shaye, D.D. and Greenwald, I. (2015). The disease-associated formin INF2/EXC-6 organizes lumen and cell outgrowth during tubulogenesis by regulating F-actin and microtubule cytoskeletons. Dev. Cell 32, 743-755. Abstract Article
Shida, T., Cueva, J.G., Xu, Z., Goodman, M.B., and Nachury, M.V. (2010). The major α-tubulin K40 acetyltransferase αTAT1 promotes rapid ciliogenesis and efficient mechanosensation. Proc. Natl. Acad. Sci. U. S. A. 107, 21517-21522. Abstract Article
Siddiqui, S. S., Aamodt, E., Rastinejad, F., and Culotti, J. (1989). Anti-tubulin monoclonal antibodies that bind to specific neurons in Caenorhabditis elegans. J. Neurosci. 9, 2963-2972. Abstract Article
Silva, M., Morsci, N., Nguyen, K.C., Rizvi, A., Rongo, C., Hall, D.H., and Barr, M.M. (2017). Cell-specific α-tubulin isotype regulates ciliary microtubule ultrastructure, intraflagellar transport, and extracellular vesicle biology. Curr. Biol. 27, 968-980 Abstract Article
Simmer, F., Moorman, C., van der Linden, A.M., Kuijk, E., van den Berghe, P.V., Kamath, R.S., Fraser, A.G., Ahringer, J., and Plasterk, R.H. (2003). Genome-wide RNAi of C. elegans using the hypersensitive rrf-3 strain reveals novel gene functions. PLoS Biol. 1, e12. Abstract Article
Singh, P., Rathinasamy, K., Mohan, R., and Panda, D. (2008). Microtubule assembly dynamics: an attractive target for anticancer drugs. IUBMB Life 60, 368-375. Abstract Article
Springer, W., Hoppe, T., Schmidt, E., and Baumeister, R. (2005). A Caenorhabditis elegans Parkin mutant with altered solubility couples α-synuclein aggregation to proteotoxic stress. Hum. Mol. Genet. 14, 3407-3423. Abstract Article
Srayko, M., Buster, D.W., Bazirgan, O.A., McNally, F.J., and Mains, P.E. (2000). MEI-1/MEI-2 katanin-like microtubule severing activity is required for Caenorhabditis elegans meiosis. Genes Dev. 14, 1072-1084. Abstract Article
Srayko, M., Kaya, A., Stamford, J., and Hyman, A.A. (2005). Identification and characterization of factors required for microtubule growth and nucleation in the early C. elegans embryo. Dev. Cell 9, 223-236. Abstract Article
Srayko, M., O'Toole E.T., Hyman, A.A., and Müller-Reichert, T. (2006). Katanin disrupts the microtubule lattice and increases polymer number in C. elegans meiosis. Curr. Biol. 16, 1944-1949. Abstract Article
Srayko, M., Quintin, S., Schwager, A., and Hyman, A.A. (2003). Caenorhabditis elegans TAC-1 and ZYG-9 form a complex that is essential for long astral and spindle microtubules. Curr. Biol. 13, 1506-1511. Abstract Article
Srivatsa, S., Parthasarathy, S., Molnár, Z., and Tarabykin, V. (2015). Sip1 downstream effector ninein controls neocortical axonal growth, ipsilateral branching, and microtubule growth and stability. Neuron. 85, 998-1012. Abstract Article
Strome, S., Powers, J., Dunn, M., Reese, K., Malone, C.J., White, J., Seydoux, G., and Saxton, W. (2001). Spindle dynamics and the role of γ-tubulin in early Caenorhabditis elegans embryos. Mol. Biol. Cell 12, 1751-1764. Abstract Article
Stubenvoll, M.D., Medley, J.C., Irwin, M., Song, M.H., (2016) ATX-2, the C. elegans ortholog of human ataxin-2, regulates centrosome size and microtubule dynamics. PLoS Genet. 12, e1006370. Abstract Article
Suprenant, K.A., Tuxhorn, J.A., Daggett, MA., Ahrens, D.P., Hostetler, A., Palange, J.M., VanWinkle, C.E., and Livingston, B.T. (2000). Conservation of the WD-repeat, microtubule-binding protein, EMAP, in sea urchins, humans, and the nematode C. elegans. Dev. Genes Evol. 210, 2-10. Abstract Article
Sze, J.Y., Liu, Y. and Ruvkun, G. (1997). VP16-activation of the C. elegans neural specification transcription factor UNC-86 suppresses mutations in downstream genes and causes defects in neural migration and axon outgrowth. Development 129, 3901-3911. Abstract Article
Teixidó-Travesa, N., Roig, J., and Lüders, J. (2012). The where, when and how of microtubule nucleation—one ring to rule them all. J. Cell Sci. 125, 4445-4456. Abstract Article
Tien, N.W., Wu, G H., Hsu, C.C., Chang, C.Y., and Wagner, O.I. (2010). Tau/PTL-1 associates with kinesin-3 KIF1A/UNC-104 and affects the motor's motility characteristics in C. elegans neurons. Neurobiol. Dis. 43, 495-506. Abstract Article
Topalidou, I., Keller, C., Kalebic, N., Hguyen, K.C.Q., Somhegyi, H., Politi, K.A., Heppenstall, P., Hall, D.H., and Chalfie, M. (2012). Genetically separable functions of the MEC-17 tubulin acetyltransferase affect microtubule organization. Curr. Biol. 22, 1057-1065. Abstract Article
Toya, M., Iida, Y., and Sugimoto, A. (2010). Imaging of mitotic spindle dynamics in Caenorhabditis elegans embryos. Methods Cell Biol. 97, 359-372. Abstract Article
Tsai, M.C., and Ahringer, J. (2007). Microtubules are involved in anterior-posterior axis formation in C. elegans embryos. J. Cell Biol. 179, 397-402. Abstract Article
Updike, D.L., and Strome, S. (2009). A genomewide RNAi screen for genes that affect the stability, distribution and function of P granules in Caenorhabditis elegans. Genetics 183, 1397-1419. Abstract Article
Wallenfang, M. R., and Seydoux, G. (2000). Polarization of the anterior-posterior axis of C. elegans is a microtubule-directed process. Nature 408, 89-92. Abstract Article
Wang, S., Wu, D., Quintin, S., Green, R.A., Cheerambathur, D.K., Ochoa, S.D., Desai, A., and Oegema K. (2015). NOCA-1 functions with γ-tubulin and in parallel to Patronin to assemble non-centrosomal microtubule arrays in C. elegans. eLife. 4, e08649. Abstract Article
Wilson, P.G., and Borisy, G.G. (1997). Evolution of the multi-tubulin hypothesis. Bioessays 19, 451-454. Abstract Article
*Edited by Michel Labouesse Last revised February 14, 2018. Published in its final form August 4, 2018. This chapter should be cited as: Hurd D.D. Tubulins in C. elegans. (August 4, 2018), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.182.1, http://www.wormbook.org.
Copyright: © 2018 Daryl D. Hurd. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
§To whom correspondence should be addressed. E-mail: dhurd@sjfc.edu
 All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.
All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.