Embryo series courtesy of Einhard Schierenberg
Embryo series courtesy of Einhard SchierenbergTable of Contents
Abstract
Transforming Growth Factor-β (TGF-β) superfamily ligands regulate many aspects of cell identity, function, and survival in multicellular animals. Genes encoding five TGF-β family members are present in the genome of C. elegans. Two of the ligands, DBL-1 and DAF-7, signal through a canonical receptor-Smad signaling pathway; while a third ligand, UNC-129, interacts with a noncanonical signaling pathway. No function has yet been associated with the remaining two ligands. Here we summarize these signaling pathways and their biological functions.
The Transforming Growth Factor-β (TGF-β) superfamily of intercellular signaling molecules constitutes a significant, ancestral means of cell-to-cell communication in eukaryotic animals (De Robertis, 2008). TGF-β signaling is required during development and homeostasis for several vital processes, including patterning the embryonic body plan and later specifying and maintaining cell identities (Wu and Hill, 2009). Alteration of TGF-β signaling causes birth defects, disorders, and diseases in humans (Padua and Massagué, 2009; Wu and Hill, 2009).
Seminal contributions based on research in the C. elegans system have proven instrumental in understanding the TGF-β signaling pathway, its transcriptional targets, and its regulators, as we discuss here. In comparison to the 30+ human TGF-β members, C. elegans has only five TGF-β ligands with non-redundant, non-lethal functions (Patterson and Padgett, 2000). TGF-β signaling pathways are highly conserved at the molecular and functional level, as TGF-β pathway components and regulators identified in C. elegans have been shown to be used by higher organisms in a related fashion (Table 1).
Table 1. TGF-β superfamily signaling is conserved in eukaryotes.
| Component | C. elegans gene name | Drosophila gene name | Human gene name | Molecule or family |
| Ligand | dbl-1 | dpp | BMP5 | Transforming Growth Factor-β |
| daf-7 | dawdle | GDF11 | ||
| unc-129 | - | - | ||
| tig-2 | gbb | BMP8 | ||
| tig-3 | dActivin | BMP2 | ||
| Type I receptor | sma-6 | tkv | BMPRIB | ser/thr kinase receptor |
| daf-1 | babo | TGF-βRI | ||
| Type II receptor | daf-4 | put | ACTRIIB | |
| R-Smad | sma-2 | Mad | Smad1 | Smad |
| sma-3 | Mad | Smad5 | ||
| daf-8 | Smox | Smad8 | ||
| daf-14 | Smox | Smad2 | ||
| Co-Smad | sma-4 | Medea | Smad4 | |
| daf-3 | Medea | Smad4 | ||
| I-Smad | tag-68 | Dad | Smad6 | |
| Transcription Factors | sma-9 | schnurri | Shn1/Shn2/Shn3 | Zn finger protein |
| lin-31 | fd96C | FoxB1 | HNF3/forkhead family | |
| mab-31 | - | - | ||
| daf-5 | - | Sno/Ski | Proline-rich protein | |
| daf-12 | Hr96 | Vitamin D receptor | nuclear hormone receptor | |
| Extracellular Regulators | lon-1 | - | - | CRISP PR-protein |
| lon-2 | Dally, Dally-like | GPC1-GPC6 | glypican | |
| sma-10 | kekkon | LRIG | leucine rich and immunoglobulin-like domains protein | |
| crm-1 | CRIM | CRIM/CHRD/chordin | cysteine-rich | |
| drag-1 | - | RGMA, RGMB | repulsive guidance molecule (RGM) | |
| adt-2 | - | - | ADAMTS |
This large family of secreted peptide growth factors in metazoans comprises a number of subfamilies, including TGF-β (the founding member of the family), activin, Nodal, Growth and Differentiation Factors (GDFs), and the Bone Morphogenetic Proteins (BMPs) (Herpin et al., 2004). The canonical TGF-β signal transduction pathway is composed of two pairs of transmembrane serine/threonine kinase receptors (type I and type II) and two or three intracellular Smad signal transducers (ten Dijke and Hill, 2004). Upon TGF-β ligand binding, the type I and type II receptors are recruited into a heterotetrameric complex and the type II receptors phosphorylate the type I receptors' conserved GS (glycine-serine) domain. The type I receptor kinase is thereby activated, and phosphorylates Smads of the R-Smad (receptor-regulated) subfamily. R-Smads form heteromeric complexes with Co-Smads (common mediator Smads) and accumulate in the nucleus to regulate gene transcription (Massagué et al., 2005). A third group of Smads, I-Smads (inhibitory Smads), usually inhibits pathway signaling by interfering with productive molecular interactions of the other Smads (Itoh et al., 2000). Smads contain two regions of homology, the N-terminal MAD homology 1 (MH1) domain that binds DNA and the C-terminal MH2 domain that participates in intermolecular interactions and regulates transcription (Wrana and Attisano, 1996).
In C. elegans, five TGF-β-related genes have been identified by sequence homology: dbl-1 (p.k.a. cet-1), daf-7, unc-129, tig-2, and tig-3 (Table 1). No biological roles have yet been described for tig-2 or tig-3 (see previous version of this chapter, TGF-β signaling; Suzuki et al., 1999). The unc-129 signaling pathway acts through a netrin receptor, UNC-5, rather than known TGF-β-related receptors or Smads (MacNeil et al., 2009). Conventional signaling pathways have been identified that act downstream of dbl-1 (Figures 1A-C, 2) and daf-7 (Figures 1D, 3). Many of the components of these two pathways were initially identified in forward genetic screens. Subsequent work using RNA interference (RNAi), reverse genetics, yeast two-hybrid assays, and DNA microarray analyses have continued to identify components, modifiers, and targets of these pathways and to uncover new biological roles for existing pathways and components. In addition to the genes described below, there is one potential I-Smad gene in the genome, tag-68, whose function has not yet been determined (Newfeld et al., 1999).
|
Figure 1. Phenotypes of DBL-1 and DAF-7 TGF-β-related pathways. A) An animal lacking functional DBL-1 is smaller than wild type. B) Wild-type animal. C) An animal overexpressing functional DBL-1 is longer than wild type. D) Animals lacking functional DAF-7 enter dauer at 25 °C. All animals were staged and imaged at a similar age at 25 °C (when animals in A-C were 24-hour adults). The anterior is left; dorsal is up. Representative individuals are shown. All images were taken at the same magnification and treated identically.
The DBL-1 pathway was first associated with two mutant phenotypes, small body size (Sma) and male tail abnormal morphology (Mab), and is also known as the Sma/Mab pathway (Padgett et al., 1998; Savage et al., 1996). Subsequent studies have shown that the DBL-1 pathway also regulates innate immunity, reproductive aging, mesodermal patterning, and chemosensation (Almedom et al., 2009; Foehr et al., 2006; Luo et al., 2009; Mallo et al., 2002; Vashlishan et al., 2008). In spite of these numerous functions, DBL-1 pathway activity is not essential for viability, a feature that has greatly facilitated genetic manipulations (Morita et al., 1999; Suzuki et al., 1999).
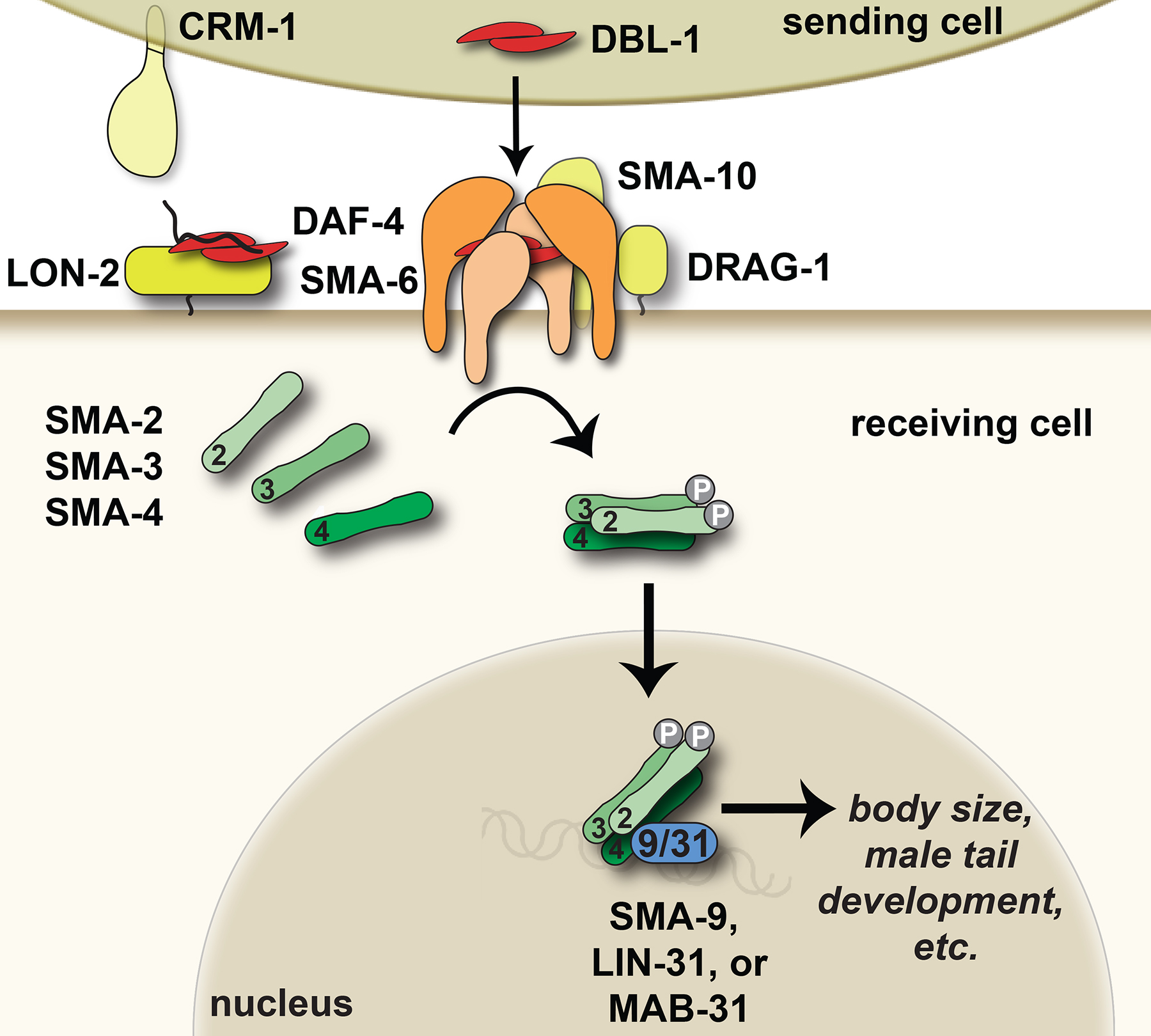
The core components of the pathway are the ligand DBL-1 (Dpp and BMP-like-1) (Morita et al., 1999; Suzuki et al., 1999), type II receptor DAF-4 (Estevez et al., 1993), type I receptor SMA-6 (Krishna et al., 1999), R-Smads SMA-2 and SMA-3, and Co-Smad SMA-4 (Figure 2) (Savage et al., 1996). Overall, these components show the greatest degree of homology to BMP subfamily ligands and signal transducers (Table 1) (Patterson and Padgett, 2000). In addition to the core components, modulators of the pathway regulate signal strength, some of them in a tissue-specific manner. Extracellular or transmembrane regulators include LON-1 (Maduzia et al., 2002; Morita et al., 2002), LON-2 (Gumienny et al., 2007), SMA-10 (Gumienny et al., 2010), CRM-1 (Fung et al., 2007), DRAG-1 (Tian et al., 2010), and ADT-2 (see section 2.7) (Fernando et al., 2011). Transcription factors that regulate signaling downstream of the receptors include SMA-9 (Liang et al., 2003), LIN-31/forkhead (Baird and Ellazar, 1999), and MAB-31 (see section 2.8) (Wong et al., 2010). dbl-1 expression is itself regulated in motor neurons by transcription factor UNC-3 (Kratsios et al., 2011).
|
Figure 2. The Sma/Mab TGF-β-related pathway. DBL-1 regulates body size, male tail morphogenesis, cell lineage decisions, and innate immune responses. The DBL-1 signal (red) is received by a heterotetrameric receptor composed of two SMA-6 type I receptor and two DAF-4 type II receptor subunits (light and dark orange, respectively). Extracellular regulators (yellows) of DBL-1 signaling include CRM-1, which acts at the sending cell membrane or in the extracellular space to promote DBL-1 signaling, SMA-10 and DRAG-1, which promote DBL-1 signaling at the membrane of the receiving cell, and LON-2/glypican, which inhibits DBL-1 signaling at the receiving cell. The DBL-1 signal is transduced by SMA-2, SMA-3, and SMA-4 Smads (greens). Transcription factors (blue) that act with Smads to carry out DBL-1-mediated responses include SMA-9/Schnurri, LIN-31/forkhead, and MAB-31. Cartoons are based on C. elegans or other published homologous protein structures and functions. See text for more details.
Sma mutant alleles were originally identified in Sydney Brenner's classic screen by their relatively proportional reduction in size (Brenner, 1974). Later, a directed genetic screen for mutant animals with small body size expanded the collection of Sma alleles (Savage-Dunn et al., 2003). Both of these screens identified mainly components of the DBL-1 pathway, establishing this pathway as the predominant regulator of body size in C. elegans. On the other hand, conserved signaling pathways that affect body or organ size in other organisms, including insulin signaling, TOR kinase, and Hippo-Warts pathways, appear to play a more minor role in size regulation in C. elegans and act in parallel with the DBL-1 pathway (Cai et al., 2009; Halder and Johnson, 2010; Jones et al., 2009; Oldham et al., 2000; Schmelzle and Hall, 2000). MAPK signaling (through SMA-5/MAPK) also affects body size in C. elegans in a manner independent from DBL-1 signaling (Watanabe et al., 2007; Watanabe et al., 2005). These studies suggest that DBL-1 acts synergistically with, instead of through, these other signaling pathways to regulate body size.
DBL-1 is a dose-dependent regulator of growth, since its functional loss (by mutation or RNAi) causes small body size, its overexpression results in long body size, and dbl-1(+/-) heterozygotes have an intermediate phenotype (Figure 1A-C) (Morita et al., 1999; Suzuki et al., 1999). Sma/Mab pathway mutant animals hatch as first larval stage (L1) larvae indistinguishable in size from the wild type (Savage-Dunn et al., 2000; Suzuki et al., 1999). Postembryonic growth is reduced in Sma/Mab mutant animals, resulting in relatively smaller body lengths and volumes (Hirose et al., 2003; Savage-Dunn et al., 2000). Sma/Mab mutant adults contain the same number of nuclei as wild-type adults, indicating that cell size must be smaller (Nagamatsu and Ohshima, 2004). Despite the relatively proportional appearance of Sma/Mab mutant animals, organ size measurements have shown that different organs are reduced in size to different degrees. The degree of reduction in the size of hypodermal (epidermal) seam cells and the large hypodermal syncytium hyp7 is most proportional to the degree of reduction in body size (Nagamatsu and Ohshima, 2004; Nyström et al., 2002; Wang et al., 2002). In contrast, neuronal soma size is not modulated by the DBL-1 pathway (Goldsmith et al., 2010). Studies directing expression of the receptors and Smads to particular tissues have demonstrated that the hypodermis, but not the intestine, plays a major role in regulation of body size (Inoue and Thomas, 2000a; Schulenburg et al., 2004; Wang et al., 2002; Yoshida et al., 2001). The gonad regulates body size, too, but through a mechanism independent of DBL-1 signaling (Patel et al., 2002).
What are the molecular mechanisms of cell size control targeted by DBL-1 pathway Smads? A correlation between body size and hypodermal ploidy has been described for some mutant strains (Flemming et al., 2000; Lozano et al., 2006; Morita et al., 2002; Nyström et al., 2002), but is not sufficient to account for all of the differences in size seen in mutant animals (Nagamatsu and Ohshima, 2004; Nyström et al., 2002). Expression profile analyses suggest that the DBL-1 pathway regulates body size in a complex way, in part through regulation of cuticle collagen and metabolism genes, as well as through signaling molecules such as insulin-like ligands and Hedgehog-related warthog ligands (see section 6.2) (Liang et al., 2007; Roberts et al., 2010). A cuticle collagen gene, lon-3, and an extracellular matrix protease gene, adt-2, may act downstream of or in parallel with the DBL-1 pathway to regulate body size (Suzuki et al., 2002).
The normal adult male tail ends in a mating structure containing nine bilaterally symmetrical pairs of sensory rays embedded in a cuticular fan (Sulston et al., 1980). In Sma/Mab pathway mutant males, defects in the specification of ray identities lead to the fusion of rays (4-5, 6-7, and 8-9) with varying penetrance (Baird and Ellazar, 1999; Krishna et al., 1999; Savage et al., 1996; Suzuki et al., 1999). Ray fusion in sma-6(wk7) males is preceded by ectopic formation of adherens junctions between cells of different rays (Hahn and Emmons, 2003). Ray identity specification is also abnormal, with the neurons of rays 5, 7, and 9 losing their dopaminergic fate (Lints and Emmons, 1999). Conversely, ubiquitous expression of dbl-1 driven by a heat shock promoter can induce ectopic expression of dopaminergic fate in neurons of rays 3, 4, 6, and 8 (Lints et al., 2004). To specify ray identities, DBL-1 pathway signaling regulates the expression of two transcription factors. DBL-1 pathway activity establishes dopaminergic fates in the A neurons of rays 5, 7, and 9 by blocking the activity of DMD-3/ DM (Doublesex/MAB-3) domain transcription factor (Siehr et al., 2011). DBL-1 pathway signaling is required for the expression of ceh-13/ labial Hox family transcription factor in the Bγ rectal blast cell lineage and ray complexes 5, 7, and 9, though males lacking ceh-13 still have functional copulatory structures (Stoyanov et al., 2003). This positive regulation of a labial homolog by a TGF-β superfamily member is conserved in Drosophila (Immergluck et al., 1990).
In addition to the sensory ray defects, the copulatory spicules are crumpled in mutant adult males (Baird and Ellazar, 1999; Krishna et al., 1999; Suzuki et al., 1999). These Mab defects prevent homozygous mutant males from mating successfully. A weak mutation in sma-6 or sma-4 that affects body size does not result in male ray fusion or spicule crumpling, indicating that body size is more sensitive to DBL-1 dosage than the Mab phenotypes (Krishna et al., 1999).
In a screen for factors involved in the morphogenesis of the male tail tip, an additional role for Sma/Mab components was uncovered (Nelson et al., 2011). RNAi knockdown of daf-4, sma-2 or sma-3 resulted in a pointed (leptoderan) rather than a rounded tail tip. Male tail morphogenesis was associated with nuclear localization of tagged SMA-3, where SMA-3 could direct target gene activity, but the hermaphrodite tail at the same developmental stage maintained tagged SMA-3 in the cytoplasm, consistent with the model that active DBL-1 pathway signaling is required to remodel the male tail. Whether this role also depends on apical junction remodeling, as the male tail ray studies suggest, remains to be discovered. For more information on male tail development and function, refer to Male development.
The DBL-1 pathway was implicated in innate immunity when it was shown that some genes regulated by DBL-1 signaling were also induced upon S. marcescens infection (Mallo et al., 2002; Mochii et al., 1999). DBL-1 highly regulates the expression of genes associated with mounting an effective innate immune response, including lectins, digestive enzymes like lysozymes and lipases, the PGP (P-glycoprotein) subclass of ATP-binding cassette (ABC) transporter family, and a caenacin (cnc) antimicrobial peptide gene cluster (Alper et al., 2007; Liang et al., 2007; Mallo et al., 2002; Mochii et al., 1999; Roberts et al., 2010). Functional loss of many DBL-1 core pathway genes increases sensitivity to a number of pathogens that infect broad ranges of hosts, from nematodes to humans, and include bacteria E. coli, E. faecalis, P. aeruginosa strain PA14, S. enterica, S. typhimurium strain SL1344, and Se. marcescens, and the nematophagous fungus D. coniospora (Mallo et al., 2002; Tenor and Aballay, 2008; Zugasti and Ewbank, 2009; So et al., 2011; Portal-Celhay et al., 2012). sma-3 was isolated in a screen to identify genes that when mutated lead to an enhanced susceptibility to pathogens (Esp), specifically P. aeruginosa strain PA14 (Kurz and Tan, 2004). All three Smads are required for a response to P. aeruginosa strain PA14, but for a response to D. coniospora, only sma-3 is needed, not sma-2 or sma-4 (Zugasti and Ewbank, 2009). This work suggests that different Smad pathways are used to regulate distinct sets of immune effectors to provide resistance to specific pathogens (Zhang and Zhang, 2009). DBL-1 pathway mutant animals may also have increased sensitivity to infection because they fail to effectively kill and digest pathogenic bacteria (So et al., 2011). DBL-1 receptors and Smads act in the pharynx and intestine, where they may contribute to innate immune responses (Krishna et al., 1999; Mallo et al., 2002; Patterson et al., 1997; Reece-Hoyes et al., 2007; Wang et al., 2002; Yoshida et al., 2001). This role for TGF-β signaling in immune responses appears to be conserved, as the Drosophila homolog of DBL-1, Decapentaplegic (Dpp), is upregulated upon infection by Micrococcus luteus (Irving et al., 2001), and mammalian TGF-βs also play roles in immune responses (Yoshimura et al., 2010). For more information, see Signaling in the immune response.
The DBL-1 pathway plays a minor role in determining lifespan. Under standard culture conditions, dbl-1 mutant populations have a somewhat reduced average lifespan relative to controls (Mallo et al., 2002; So et al., 2011). However, there is evidence that this reduced lifespan is due to increased susceptibility of DBL-1 pathway mutant animals to the slight pathogenic effects of the standard bacterial food strain, OP50, to which animals lacking DBL-1 are more susceptible than the wild type. Consistent with this hypothesis, dbl-1 mutant animals grown on OP50 accumulate a greater number of live bacterial cells in their intestines than wild-type animals grown under the same conditions (So et al., 2011). Furthermore, growth of dbl-1 mutant animals on heat-killed OP50 increases their survival (Mallo et al., 2002; So et al., 2011). When 5-fluoro-2′-deoxyuridine (FUdR) is added to plates to prevent DNA synthesis, thus also preventing progeny production and bacterial replication, animals lacking DBL-1 no longer have reduced survival but instead populations show moderate increases in lifespan (Luo et al., 2009; Mallo et al., 2002). These results suggest that the pathogenic effect of OP50 is obscuring a moderate lifespan-extending effect of dbl-1 inactivation. Consistent with this hypothesis, dbl-1 overexpression under these conditions reduces lifespan (Luo et al., 2009). Another proposed mechanism by which DBL-1 pathway mutations may extend lifespan is through the established lifespan extending factor dietary restriction (So et al., 2011), since expression of some digestive enzymes is positively correlated with DBL-1 pathway activity (Liang et al., 2007; Roberts et al., 2010).
In contrast to its minor effects on lifespan, the DBL-1 pathway plays a major role in the regulation of reproductive aging (Luo et al., 2010; Luo et al., 2009). As in humans, cessation of reproduction in C. elegans normally occurs in mid-adulthood. In wild-type hermaphrodites, the mean reproductive span is approximately 3.5 days, while that for dbl-1 mutant animals is more than 7 days (Luo et al., 2009). Other DBL-1 pathway alleles also permit significantly longer reproductive spans than controls (Luo et al., 2009). This result indicates that the DBL-1 pathway normally restricts reproductive span and promotes timely cessation of reproduction. Mechanistically, the DBL-1 pathway regulates reproductive aging by modulating germline and oocyte quality (Luo et al., 2010). Not only is the population of proliferating germ cells maintained later in life in sma-2 mutant animals, these cells show significantly reduced indicators of decline, including cavities, graininess, and cellularization. These healthier germ cells produce fertilizable oocytes later in life than controls. Oocytes from older sma-2 mutant hermaphrodites have visibly improved oocyte morphology, a lower frequency of chromosomal nondisjunction, and greater success in developing into viable embryos than oocytes from similarly aged wild-type animals. Using tissue-specific expression of sma-3, Luo et al. demonstrated that the DBL-1 pathway acts non-autonomously in somatic tissues to regulate reproductive aging (Luo et al., 2010). Finally, this regulation of reproductive aging is independent of the FoxO transcription factors daf-16 and pha-4 (Luo et al., 2009), which mediate DAF-7/insulin signaling and dietary restriction regulation of longevity, respectively (see section 6.2). Thus, the DBL-1 pathway uncouples the regulation of lifespan from that of reproductive aging.
In the postembryonic mesodermal lineage, the M blast cell divides to produce descendants that contribute to the sex muscles (SM fate) and to the coelomocytes (CC fate), pseudocoelomic scavenger cells (Sulston and Horvitz, 1977). In genetic screens for mutant animals with alterations in the M lineage, two sma-9/Schnurri alleles were identified (Foehr et al., 2006). These alleles cause a duplication of SMs at the expense of CCs. Surprisingly, alleles affecting core DBL-1 signaling components produce no M lineage defect. Instead, these mutant alleles can function as suppressors of the sma-9 M lineage phenotype (SOSMLP) in double mutant combinations. While SMA-9/Schnurri promotes body size signaling, this phenomenon suggests that sma-9 antagonizes DBL-1 pathway signaling to promote dorsal M lineage fates (Foehr and Liu, 2008). The SOSMLP phenotype was used to screen for novel regulators of the DBL-1 pathway. drag-1, which encodes a membrane-associated repulsive guidance molecule (RGM) protein, was isolated from this screen and identified as a positive regulator of DBL-1 activity (Tian et al., 2010). This was the first evidence that an RGM molecule regulates TGF-β signaling in vivo, though in vitro evidence in mammalian systems has shown that RGM proteins can act as co-receptors for the TGF-β member BMP6 (Andriopoulos et al., 2009; Babitt et al., 2007; Tian et al., 2010; Xia et al., 2008).
DBL-1 is expressed in and secreted from cholinergic neurons, which excite neighboring body muscles (Duerr et al., 2008; Morita et al., 1999; Suzuki et al., 1999). The DBL-1 pathway plays a clear role in how nematodes respond to excitatory acetylcholine inputs, as mutations in dbl-1, sma-6, sma-3, or sma-4 confer hypersensitivity to cholinergic agonists levamisole and nicotine, and/or the acetylcholinesterase inhibitor aldecarb (Almedom et al., 2009; Vashlishan et al., 2008). Increased DBL-1 pathway signaling results in animals that are resistant to levamisole (Gumienny laboratory, unpublished observations). Hypersensitivity of muscle cells to these drugs is caused by mutations that increase acetylcholine secretion or decrease inhibitory GABAergic synaptic signaling (Loria et al., 2004; Gracheva et al., 2006; McEwen et al., 2006). Loss of DBL-1 results in both pre- and post-synaptic defects at GABAergic synapses, suggesting that this sensitivity occurs through a defect in GABAergic signaling (Vashlishan et al., 2008). However, the molecular basis of how the DBL-1 pathway may normally promote GABA transmission is unknown.
The DBL-1 receptors SMA-6 and DAF-4 and SMA-4/Smad also appear to play a role in maintaining normal serotonin levels by acting downstream of a serotonin pathway calcium channel, UNC-2, and repressing expression of tph-1, a tryptophan hydroxylase gene required for serotonin synthesis (Estevez et al., 2004). These transmission defects may underlie the slow movement observed in dbl-1(-) animals (Maniere et al., 2011).
Another neuronal function attributed to DBL-1 is its effect on fluorescently tagged, misfolded superoxide dismutase SOD1(G85R) expressed in neurons (Wang et al., 2009). C. elegans overexpressing a human SOD1 variant that causes amyotrophic lateral sclerosis (ALS, Lou Gehrig's disease) in humans develop aggregates of this protein within the expressing neurons and develop motor defects over time. Loss of dbl-1 activity exacerbated protein aggregation and led to earlier onset of movement defects, suggesting that DBL-1 normally either prevents aggregation or promotes degradation of this misfolded protein.
DBL-1 was identified both by forward genetics using the small body size phenotype and by reverse genetics due to its homology to Drosophila Dpp and mammalian BMPs (Morita et al., 1999; Suzuki et al., 1999). DBL-1 is secreted from neurons in the head and nerve cords. Heterologous expression of dbl-1 demonstrates that the tissue source of DBL-1 is less important than the level of expression for DBL-1 signaling. This suggests that DBL-1 expression levels may normally depend on cues processed by the neurons that express DBL-1 (Savage-Dunn et al., 2011). A major target of DBL-1 for body size regulation appears to be the hypodermis, which cradles the nerve cords along their entire length. The hypodermis expresses the receptors for DBL-1, SMA-6/type I, which is pathway-specific (Krishna et al., 1999), and DAF-4/type II, which also functions in the dauer pathway (Estevez et al., 1993).
Between the TGF-β secreting cells and the receiving cells, the TGF-β does not simply diffuse to its receptors on the receiving cell, but is tightly regulated. In C. elegans, four transmembrane proteins have been identified that have tissue-specific roles in extracellular regulation of DBL-1 activity: the negative regulator LON-2/glypican and the positive regulators SMA-10/LRIG, CRM-1/CRIM, and DRAG-1/Dragon. LON-2, one of the two glypicans in C. elegans, is a heparan sulfate proteoglycan that negatively regulates DBL-1 signaling (Gumienny et al., 2007). Like Drosophila glypican Dally, LON-2 binds BMP members and thereby is proposed to regulate signaling (Gumienny et al., 2007; Kirkpatrick et al., 2006). Loss of functional heparan sulfate modifying enzymes results in small animals that may be a result of LON-2 over-repressing DBL-1, since LON-2 is required for the small phenotype of the heparan sulfate enzyme mutant animals (Gumienny et al., 2007).
SMA-10 is a positive regulator of DBL-1 signaling. It is member of the leucine-rich repeats and immunoglobulin-like domains (LRIG or LRRIG) family (Gumienny et al., 2010). This conserved transmembrane protein binds to both SMA-6/type I and DAF-4/type II receptors in cell culture assays, as does one of its mammalian homologs, LRIG1.
CRM-1, a member of the cysteine-rich motor neuron protein (CRIM) family, also acts in the intercellular milieu to facilitate DBL-1 signaling (Fung et al., 2007). Although CRM-1 is expressed in the same ventral cord neurons as DBL-1, this transmembrane protein functions in a cell non-autonomous way to promote DBL-1 signaling in the hypodermis. Post-translational processing or alternative splicing to make soluble CRM-1 isoforms may account for this activity. In vertebrate systems, CRIM has also been shown to bind BMPs, but reduces BMP processing and secretion, thereby antagonizing BMP activity (Wilkinson et al., 2003).
The third extracellular positive regulator of DBL-1 signaling is DRAG-1, the sole member of the repulsive guidance molecule (RGM) family in C. elegans. DRAG-1 promotes normal body size and mesoderm development through the DBL-1 pathway (Tian et al., 2010). Like its conserved family members in other species, DRAG-1 is GPI-linked to the extracellular membrane surface. Other RGM members have been shown to physically bind specifically to some BMPs and both BMP receptor types (Corradini et al., 2009).
In addition to these transmembrane regulators of DBL-1 signaling, ligand activity may also be regulated through interactions with the extracellular matrix. For example, DBL-1 genetically acts with the extracellular matrix constituent UNC-52/perlecan in directing migrating distal tip cells (Merz et al., 2003). Perlecan in other systems interacts with the TGF-β regulator fibrillin-1 (Tiedemann et al., 2005). DBL-1 signaling is also positively regulated in part by ADT-2 (p.k.a. SMA-21), a member of the ADAMTS (a disintegrin and metalloprotease with thrombospondin motifs) family of secreted metalloproteases (Fernando et al., 2011). Animals lacking functional ADT-2 are smaller than normal and, in this background, expression of a DBL-1-responsive transcriptional reporter is decreased. However, ADT-2 is also required for normal fibril formation of cuticle collagens, which may affect body size in a DBL-1-independent manner (Fernando et al., 2011). ADAMTS metalloproteases assemble and degrade extracellular matrix (Colige et al., 1995; Collins-Racie et al., 2004; Fernandes et al., 2001; Kuno et al., 2000; Somerville et al., 2003; Tortorella et al., 2000; Tortorella et al., 2005; Wang et al., 2003). Mouse cells lacking ADAMTS5 have increased responsiveness to TGFβ1 treatment (Velasco et al., 2011), while geleophysic dysplasia patients with altered ADAMTSL2 show increased TGF-β activity (Le Goff et al., 2008). ADAMTSL2 binds the TGF-β inhibitor latent TGF-β-binding protein 1 (LTBP1), which does not have an identified homolog in C. elegans (Le Goff et al., 2008).
The main intracellular signal transducers for this pathway are SMA-2, SMA-3, and SMA-4 (Savage et al., 1996), which, together with Drosophila Mad, were the founding members of the Smad family of transcription factors (Sekelsky et al., 1995). SMA-2 and SMA-3 are receptor-regulated Smads (R-Smads), and SMA-4 is a Co-Smad. Smads engage in dynamic nucleocytoplasmic shuttling, with retention in the nucleus stimulated by pathway activation (Inman et al., 2002; Nicolás et al., 2004; Xiao et al., 2001; Xu et al., 2002). In C. elegans, a rescuing SMA-3:GFP fusion protein normally accumulates to high levels in the nucleus, while in sma-6 type I receptor mutant animals, the protein is more evenly partitioned between nucleus and cytoplasm (Wang et al., 2002). Nuclear accumulation is not influenced by R-Smad (SMA-2) or Co-Smad (SMA-4) depletion. SMA-2 and SMA-3 R-Smads are not redundant for body size and male tail patterning (Savage-Dunn et al., 2000), suggesting that the functional Smad complex is a heterotrimer containing two different R-Smads (SMA-2/SMA-3/SMA-4). In contrast, mounting an innate immune response against the fungus D. coniospora has provided evidence of a noncanonical DBL-1 signaling mechanism that relies on SMA-3 but not SMA-2 or SMA-4 (see section 2.3) (Zugasti and Ewbank, 2009). This result suggests that during an innate immune response, the functional Smad complex is an R-Smad homo-multimer that does not incorporate a Co-Smad. Expression of the Smads and their receptors in the hypodermis, pharynx, and intestine are involved in immune responses, while the body size regulatory aspect of Smads is localized only in the hypodermis and pharynx (Krishna et al., 1999; Schulenburg et al., 2004; Wang et al., 2002; Yoshida et al., 2001).
Another intracellular modulator of TGF-β signaling in C. elegans is KIN-29 (p.k.a. SMA-11 and SNS-8), a serine/threonine kinase that acts downstream of DBL-1 to regulate body size (Lanjuin and Sengupta, 2002; Maduzia et al., 2005). kin-29 is expressed in a number of sensory neurons, including some that express dbl-1, where it appears to affect levels of some olfactory receptors (Lanjuin and Sengupta, 2002). KIN-29 is also expressed in the hypodermis, where DBL-1 receptors and downstream players and targets reside, though its targets in the hypoderm are unknown (Maduzia et al., 2005). Body size is partially restored in kin-29 mutant animals if wild-type kin-29 is expressed in either neurons or hypodermis, but the molecular mechanism of this rescue is currently unknown.
SMA-9 is a transcription factor in the Sma/Mab pathway (Liang et al., 2003). Identified in genetic screens for small (Savage-Dunn et al., 2003) or mesodermal patterning mutant animals (Foehr et al., 2006), sma-9 encodes a large zinc finger transcription factor related to Drosophila Schnurri (Liang et al., 2003). SMA-9 is nuclear localized and is detected in most, if not all, somatic cells by immunohistochemistry. Phenotypic analyses of sma-9 indicates that it functions in a subset of Sma/Mab signaling outcomes: early but not late larval growth; ray 9 but not ray 5 and 7 morphogenesis; regulation of neurotransmitter expression in rays 5, 7, and 9; and spicule morphogenesis with reduced penetrance. In mesodermal patterning, SMA-9 acts antagonistically with the DBL-1 pathway (see section 2.5) (Foehr et al., 2006). SMA-9 is not required for DBL-1 transcriptional regulation of antimicrobial peptide gene expression following fungal infection (Zugasti and Ewbank, 2009). These complexities in function may depend on alternative splicing to encode isoforms of SMA-9 that have varying zinc finger motif numbers. This model has been demonstrated in the case of SMA-9 in male tail development, where alternative transcript processing results in a truncated SMA-9 required for male tail patterning but not for body size regulation (Yin et al., 2010).
Additional transcriptional partners have been identified that play tissue-specific roles in TGF-β signal transduction. For example, the Forkhead/Fox transcription factor LIN-31, originally identified by its role in vulval development (Miller et al., 1993), is involved in spicule morphogenesis with the Sma/Mab pathway (Baird and Ellazar, 1999). Yeast two-hybrid experiments demonstrate that LIN-31 can physically interact with SMA-3 (Wang et al., 2005). In vertebrates, FoxO and FoxH transcription factors are Smad transcriptional partners (Massagué et al., 2005), suggesting that this Smad-Fox transcriptional partnership is evolutionarily conserved. Another tissue-specific transcription partner is MAB-31. mab-31 was identified in an RNAi screen for genes required for normal male tail development (Wong et al., 2010). Loss of mab-31 function, either by RNAi or in a deletion mutant, results in ray fusions indistinguishable from the core Sma/Mab pathway mutant phenotype. MAB-31:GFP localizes to the nucleus, and its localization does not depend on SMA-6 receptor or SMA-4 Co-Smad, suggesting constitutive nuclear localization rather than nucleocytoplasmic shuttling. MAB-31 is conserved among nematodes, but shows no apparent homology to proteins in other organisms. These three transcription factors, SMA-9, LIN-31 and MAB-31, can be considered integral components of the DBL-1 pathway based on the close similarity of their mutant phenotypes to those of other DBL-1 pathway mutant animals. Other transcription factors that are required for DBL-1-regulated responses are more pleiotropic and are described below.
Body size regulation by the DBL-1 pathway may, like other TGF-β pathways, require accessory transcription factors. Smads, by themselves, interact poorly with DNA and complex with other transcription factors to mediate TGF-β responses (reviewed by Massagué et al., 2005; Schmierer and Hill, 2007). In C. elegans, RNT-1 (RUNX family transcription factor) has been identified as a transcription factor that physically interacts with SMA-4 Smad in vitro and is required for normal body size, as loss of RNT-1 function results in small animals (Ji et al., 2004). In mammals, RUNX transcription factors also interact with Smads (Hanai et al., 1999; Selvamurugan et al., 2004; Zhang et al., 2000). rnt-1 is expressed in the seam cells and the intestine, but not apparently in the hypodermis, so how RNT-1 affects hypodermally-expressed SMA-4 remains unclear (Nam et al., 2002). Double mutant combinations with rnt-1 and sma-2, sma-3, sma-4, or sma-6 result in animals smaller than animals harboring a single mutant allele, suggesting that rnt-1 acts at least partly in parallel to DBL-1 signaling for body size, possibly through different transcriptional targets or tissues (Ji et al., 2004). This SMA-4—RNT-1 interaction appears to be inhibited by another transcription factor, BRO-1 (brother (Drosophila transcription factor partner) homolog), which competes with DBL-1 signaling by interacting with RNT-1 in a complex with DNA (Shim and Lee, 2008).
In the male tail rays, DBL-1 promotes a dopaminergic fate in neurons of rays 5, 7, and 9 (Lints and Emmons, 1999). Regulation of this fate requires genetic interaction with the Hox gene egl-5 and the DM domain transcription factor gene mab-23 (Lints and Emmons, 1999, 2002). EGL-5 is required for rays 3-6 to be competent to express the dopaminergic fate in response to DBL-1 signaling (Lints and Emmons, 1999). On the other hand, DBL-1 induction of dopaminergic fate in ray 5 requires suppression of mab-23 activity (Lints and Emmons, 2002). MAB-23 normally functions to repress dopaminergic fate, since in mab-23 mutant adult males, rays 1, 3, and 6 ectopically express dopaminergic fate (Lints and Emmons, 2002).
Another gene that acts downstream of the DBL-1 pathway in specification of ray identity is mab-21, which encodes a novel, conserved protein (Chow et al., 1995; Morita et al., 1999). mab-21 is required in the structural cell of ray 6 to specify ray 6 identity, and its Mab phenotype is epistatic to that of the DBL-1 pathway (Ho et al., 2001; Morita et al., 1999). In C. elegans, SIN-3, a histone deacetylase complex subunit, associates with MAB-21 (Choy et al., 2007). Interestingly, vertebrate Mab21 and Sin3 both physically interact with Smads and modulate BMP signaling (Baldessari et al., 2004). These results support a model whereby a complex of Smads, MAB-21, and SIN-3 repress transcription of TGF-β target genes by recruiting a histone deacetylation complex to repress transcription of targeted genes (Inoue and Imamura, 2008).
Downstream transcriptional targets of the DBL-1 pathway have been determined using microarray analyses. Liang et al. compared expression profiles of dbl-1 and sma-9 mutant populations to the wild type at the L2 stage (Liang et al., 2007); Roberts et al. compared sma-6 mutant populations with dbl-1(++) overexpressing animals at the L4 stage (Roberts et al., 2010); Luo et al. compared sma-2 and sma-4 mutant populations to wild type at the L4 stage (Luo et al., 2010). All three groups identified similar target gene categories, including cuticle collagens, hedgehog-related genes, metabolism genes, and innate immunity genes. These results are similar to those obtained in a smaller scale arrayed cDNA hybridization experiment (Mochii et al., 1999). Differences in the specific genes identified in these experiments may be due to stage-specific regulation of target genes. In addition to the somatic targets of the pathway, germline genes showed significant decreases in expression levels (Liang et al., 2007; Luo et al., 2010; Roberts et al., 2010). By performing expression profiles on isolated unfertilized oocytes, Luo et al. found germline-specific sma-2 target genes that have no overlap with the target genes identified in the L4 stage (Luo et al., 2010). One of the functions of these target genes is to mediate the regulation of reproductive aging (see section 2.4).
The DAF-7 pathway was first elucidated by its regulation of the dauer/continuous development switch. The core components of this pathway are: the ligand DAF-7 (Ren et al., 1996), type II receptor DAF-4 (Estevez et al., 1993), type I receptor DAF-1 (Georgi et al., 1990), R-Smads DAF-8 (Park et al., 2010) and DAF-14 (Inoue and Thomas, 2000a), and antagonistic Co-Smad DAF-3 (Patterson et al., 1997) (Figure 3). The Sno/Ski homolog DAF-5 acts downstream of the pathway and binds DAF-3/Co-Smad, an interaction that is conserved in other systems (da Graca et al., 2004; Inoue and Imamura, 2008; Tewari et al., 2004). DAF-7 is expressed only in the pair of ASI sensory neurons (Ren et al., 1996; Schackwitz et al., 1996), while other pathway components have widespread expression. DAF-7 pathway components are in general quite highly diverged from TGF-β signaling components in other species, but have more sequence features in common with TGF-β/Activin/Nodal/GDF pathway components than with BMP components (see previous version of this chapter, TGF-β signaling).
|
Figure 3. The dauer TGF-β-related pathway. DAF-7 promotes continuous, non-dauer development. The DAF-7 signal (red) is received by a heterotetrameric receptor composed of two DAF-1 type I receptor and two DAF-4 type II receptor subunits (light and dark orange, respectively). BRA-1/BMP receptor associated protein (BRAM) (yellow) is a negative intracellular regulator of DAF-1. The DAF-7 signal is transduced by DAF-8 and DAF-14 Smads (light greens). These components, when activated, inhibit the functions of DAF-3/Co-Smad (dark green) and DAF-5/Sno/Ski (indigo), which promote dauer development. The Smad/Ski model shown is not definitive. Cartoons are based on C. elegans or other published homologous protein structures and functions. See text for more details.
The dauer larva is an alternative third larval stage specialized for survival under harsh environmental conditions (high population density, low food availability, high temperature) (Cassada and Russell, 1975; Golden and Riddle, 1984a, Golden and Riddle 1984b). The decision to enter the dauer stage is made in the L1 and L2 predauer stages (Swanson and Riddle, 1981). When dauer larvae sense more favorable environments, they exit dauer and resume development. Large-scale genetic screens were conducted for mutations that affect the decision to enter dauer (Golden and Riddle, 1984b; Malone and Thomas, 1994; Riddle et al., 1981). The mutant strains isolated were either dauer-constitutive (Daf-c: constitutively entering dauer inappropriately under favorable environmental conditions) or dauer-defective (Daf-d: defective in entering dauer under adverse conditions). Genetic epistasis and detailed phenotypic analyses revealed three dauer gene classes (Thomas et al., 1993). Subsequent molecular analyses established that the genes in each class encode components of three different signaling pathways: a TGF-β-related pathway, an insulin-related pathway, and a cyclic nucleotide pathway (see Dauer). Animals defective in the core components of the daf-7/TGF-β dauer pathway are Daf-c. Mutations in the downstream genes daf-3/Co-Smad and daf-5/Sno/Ski cause Daf-d phenotypes that suppress the Daf-c phenotypes of the upstream components (Figure 3) (Thomas et al., 1993).
DAF-7 pathway mutant phenotypes are temperature sensitive, with increased penetrance of dauer at 25°C than at 15°C (Golden and Riddle, 1984b). Temperature modulates the animal's response to other dauer-inducing cues, pheromone and food shortage. daf-7/TGF-β expression is responsive to changes in temperature, pheromone, and food (Ren et al., 1996; Schackwitz et al., 1996). However, daf-3 and daf-5 mutant alleles suppress the daf-7 pathway mutant Daf-c phenotype at 25°C but enhance it at 27°C, and daf-3 or daf-5 mutant animals retain pheromone responsiveness, suggesting that DAF-7 pathway signaling, pheromone levels, and temperature are three different cues the organism must integrate in its decision to enter dauer (Ailion and Thomas, 2000; Ren et al., 1996; Thomas et al., 1993). For more information, refer to the WormBook chapter Dauer.
Both dauer entry and exit decisions appear to require DAF-7. daf-7 expression decreases with dauering and increases during dauer exit induced by presence of food or a shift to a permissive temperature (Ren et al., 1996; Schackwitz et al., 1996). Shifting dauers with a wild-type background to a permissive temperature or from high to low pheromone concentration in the presence of food recovers dauers, but dauer recovery is greatly reduced in daf-7 mutant animals (Ren et al., 1996). Dauer recovery of daf-7 mutants is further delayed or prevented when the nuclear hormone receptor NHR-69 or Notch LIN-12d is lacking, and is enhanced at 25 oC by muscarinic agonist neurotransmitters or if the Notch pathway (GLP-1 or LAG-2) or insulin INS-18 is not functional (Tissenbaum et al., 2000; Ouellet et al., 2008; Park et al., 2012b). These results suggest that DAF-7 signaling is an important part of recovering from dauer as well as bypassing dauer in reproductively favorable environments.
In adult nematodes, the DAF-7 pathway responds to environmental signals by modulating energy consumption. In particular, daf-7, daf-4, and daf-1 mutant animals grown under favorable environmental conditions accumulate more intestinal fat than wild type, in spite of reduced feeding (Greer et al., 2008). These metabolic changes are similar to those seen in dauers or in other nematodes grown under unfavorable conditions. Like the Daf-c phenotype, these metabolic phenotypes are dependent on daf-3/Smad (Greer et al., 2008). egl-4/cGMP-dependent protein kinase may act upstream of daf-7 to promote fat deposition in the intestine (Raizen et al., 2006). To identify specific effectors of the daf-7(-) fat accumulation phenotype, a suppressor screen was performed. It identified a gain-of-function allele of egl-30, which encodes an α subunit of a heterotrimeric G protein that promotes acetylcholine secretion (Lackner et al., 1999; Miller et al., 1999; Greer et al., 2008). How this fat accumulation suppressor interacts with the DAF-7 pathway is undetermined. In addition, loss of two metabotropic glutamate receptors, MGL-1 and MGL-3, specifically eliminated the increased fat phenotype of daf-7 mutant animals. These results are consistent with a model where DAF-7 signaling during reproductively favorable conditions inhibits DAF-3 activity and permits appropriate fat deposition. In an unfavorable environment, DAF-7 is inactivated, thereby activating DAF-3 and increasing fat storage levels through MGL-1 and MGL-3 metabotropic receptors (Greer et al., 2008). For more information, refer to the WormBook chapter Obesity and the regulation of fat metabolism.
In addition to the Daf-c and fat storage phenotypes, animals lacking DAF-7 display behavioral defects in egg laying, roaming, quiescence, and feeding. The egg-laying defect is a partial retention of embryos in the uterus, reminiscent of the egg retention that occurs under low food conditions. Like other phenotypes of DAF-7 pathway mutant strains, the egg-laying defect is suppressed by mutations in daf-3/Co-Smad and daf-5/Sno/Ski (Trent et al., 1983). Mutations in two other genes required for egg laying, egl-4 and egl-32, are also suppressed by daf-3 and daf-5 mutations, indicating a possible interaction with this pathway. egl-4/cGMP-dependent protein kinase has additional phenotypes that overlap with those of both dbl-1 and daf-7, including body size regulation (see section 6.2), fat accumulation (see section 3.2), and chemosensation (see section 6.2) (Fujiwara et al., 2002; Hirose et al., 2003; L'Etoile et al., 2002). egl-32, which is uncloned, is also required for normal egg laying and chemosensation, though the molecular basis of these phenotypes and whether they are DAF-7 dependent is unknown (Daniels et al., 2000; McGovern et al., 2007).
Like other animals, C. elegans divide their time between alternative behavioral states of inactive dwelling and active roaming, with this allocation responsive to food availability (see Chemosensation in C. elegans). During dwelling, which occurs 80% of the time under normal laboratory conditions, animals move slowly, alternating between forward and backward movement, with little net displacement. In roaming, animals move in a more continuous forward direction. When food resources are low or poor, the time allocated to roaming will increase (Fujiwara et al., 2002). daf-7 mutants, however, are defective in triggering roaming behavior in response to unfavorable food conditions (Ben Arous et al., 2009). Another distinct behavioral response to food is quiescence, defined as the complete cessation of movement and feeding for periods of at least 10 seconds (You et al., 2008). Quiescence is not normally seen with standard laboratory food (OP50 bacteria), but can be triggered by feeding HB101 bacteria, which is considered a higher quality food (You et al., 2008). This quiescence may be analogous to satiety in mammals. The induction of quiescence is defective in daf-7, daf-1, and daf-4 mutant animals, as well as in insulin signaling defective animals (Ren et al., 1996; You et al., 2008). Furthermore, genetic interactions suggest that both DAF-7 and insulin pathways function via the cGMP-dependent protein kinase EGL-4 to induce quiescence.
Another behavioral alteration described in animals defective in the DAF-7 pathway is the clumpy phenotype (Thomas et al., 1993). These mutant animals tend to aggregate on a bacterial lawn, in contrast to the dispersed localization of the standard laboratory strain N2. In natural isolates of C. elegans, these two alternate behaviors are observed and have been attributed to polymorphisms in the npr-1 neuropeptide receptor gene (de Bono and Bargmann, 1998). This aggregation behavior is proposed to be a way to avoid hyperoxia, as local oxygen levels are decreased within aggregations (Gray et al., 2004). Like npr-1-defective animals, animals lacking DAF-7 also display a robust avoidance of hyperoxia and aggregate in the presence of food, unlike N2 wild-type animals, which aggregate to avoid hyperoxia only in the absence of food (Chang et al., 2006). Since npr-1 and daf-7 act in different cell types, however, they are believed to act in parallel in a distributed neuronal network regulating these behaviors.
Animals lacking a functional DAF-7 pathway have reduced feeding rates as measured by pharyngeal pumping (Greer et al., 2008). These reductions are similar to those seen in wild-type nematodes treated exogenously with the neurotransmitters tyramine or octopamine (Horvitz et al., 1982). By blocking the tyramine or octopamine biosynthetic pathways or signal transduction pathways in daf-7 mutant animals, Greer et al. showed that the reduced pumping in these animals depends on these neurotransmitters (Greer et al., 2008). The dauer, egg-laying and fat retention phenotypes, on the other hand, were unaltered by elimination of tyramine or octopamine signaling and therefore act through a different mechanism than DAF-7-regulated pharyngeal pumping. These results suggest that DAF-7 is an environmental sensor to determine not only the dauer decision, but also to modulate the animal's behaviors to maintain energy balance at other stages. For more information on the regulation of these behaviors, refer to the WormBook chapters Egg-laying, Neuropeptides, Biogenic amine neurotransmitters, Acetylcholine, and GABA.
The output of DAF-7 pathway signaling is integrated with insulin signaling in the regulation of dauer entry and exit. Early reports suggested that the functions of these two pathways diverged in adulthood, with insulin signaling but not TGF-β signaling regulating longevity (Kenyon et al., 1993; Larsen et al., 1995). It has subsequently been shown, however, that matricide masked a long-lived phenotype of DAF-7 pathway mutant strains (Shaw et al., 2007). This death of adult hermaphrodites is a consequence of embryos hatching internally and is another effect of the daf-7 mutant egg-laying defect. When lifespan assays were conducted on FudR to prevent embryonic development, a significant increase in lifespan was observed in daf-7, daf-1, daf-4, daf-8, and daf-14 mutant strains relative to wild type (Shaw et al., 2007).
The lifespan extension of daf-7 mutant populations was dependent upon daf-16/FoxO, the downstream transcription factor in the insulin pathway, suggesting that the TGF-β pathway regulates longevity upstream of and via the insulin pathway (Shaw et al., 2007). Consistent with this hypothesis, the expression profiles of DAF-7 and insulin mutants are highly similar to each other. Furthermore, the insulin-like ligands ins-7 and ins-18 are transcriptionally regulated by the DAF-7 pathway, providing a mechanism for regulation of the insulin pathway (Liu et al., 2004; Shaw et al., 2007; Narasimhan et al., 2011).
A negative regulatory influence of the DAF-7 pathway on the germline proliferative zone was suggested by Park et al. (Park et al., 2010). Based on their studies, they concluded that DAF-8/R-Smad expressed in the distal tip cell negatively regulates the expression of lag-2, which encodes the DTC-expressed ligand for GLP-1/Notch, and thereby negatively regulates the proliferative pool of germ cells. This role for DAF-8 appeared to be independent of DAF-3 and DAF-5 (Park et al., 2010).
However, in screens to identify genes involved in germline proliferation and differentiation, daf-7, daf-1, and daf-4 were identified as positive regulators of the proliferative zone (Dalfó et al., 2012). DAF-7 expressed from the ASI environment-sensing cells promotes the larval accumulation of germline progenitor cells in response to reproductively favorable environmental conditions. To coordinate this response, DAF-7 requires the activity of its receptor DAF-1 in the distal tip cells, which constitute the niche for germline stem cells. DAF-3, DAF-5, DAF-8, and likely DAF-14 are also expressed in the distal tip cells (Patterson et al., 1997; Inoue and Thomas, 2000a; Park et al., 2010; da Graca et al., 2004). DAF-3 and DAF-5 limit DAF-7-mediated expansion of the proliferative zone, while DAF-8 and DAF-14, like DAF-7 and its receptors, promote expansion (Dalfó et al., 2012). Additional results support the idea that this pathway acts to influence the proliferation versus differentiation decision of larval germ cells (as opposed to influencing the cell cycle rate), but in a GLP-1/Notch-independent manner. Though currently not reconciled, the opposite germline phenotypes observed by these two groups may be independent and may be controlled in different ways by DAF-7 pathway components. DAF-7's positive regulation of larval proliferative zone expansion is also independent of DAF-12/nuclear hormone receptor, DAF-18/PTEN, or DAF-16/FOXO, which are required for the DAF-7 response to dauering cues (DAF-12) and life-span regulation (DAF-18 and DAF-16) (see above) (Dalfó et al., 2012). These results suggest that DAF-7 influences larval germline progenitors through an unidentified downstream pathway.
DAF-7/TGF-β-related ligand is expressed in the pair of ASI sensory neurons under favorable environmental conditions and is repressed under adverse conditions, such as increased population density and low food availability, leading to appropriate responses to these conditions (Ren et al., 1996; Schackwitz et al., 1996). Animals lacking functional DAF-7 are Daf-c, indicating that this TGF-β superfamily member has a role in promoting continuous, non-dauer development. Genetic analysis of dauer formation in the closely related free-living nematode Caenorhabditis briggsae revealed similar pathways involved in dauer regulation (Inoue et al., 2007). Specifically, the C. briggsae daf-4 phenotype is Daf-c, while the C. briggsae daf-3 phenotype is Daf-d.
DAF-7 orthologs have been identified in several species of parasitic nematodes, but their roles in responding to environmental cues appear to be diverged (Brand et al., 2005; Crook et al., 2005; Freitas and Arasu, 2005; Gomez-Escobar et al., 2000; McSorley et al., 2009). Not only does Parastrongyloides trichosuri daf-7 fail to rescue C. elegans daf-7 mutant phenotypes, its expression is different than C. elegans daf-7 (Crook et al., 2010). In contrast with C. elegans daf-7 (Ren et al., 1996), which is expressed in early larval stages but is downregulated upon entry into dauer, expression of parasitic nematode daf-7 homologs increases during the infectious diapause state (Viney et al., 2005).
The type I and type II receptors for DAF-7 are encoded by daf-1 and daf-4, respectively (Estevez et al., 1993; Georgi et al., 1990). The molecular cloning of daf-4 was pivotal in the recognition of this pathway as a TGF-β signaling pathway, since DAF-4 showed homology to the newly identified TGF-β type II receptors. (The cloning of daf-1 preceded this time, so its homology was not recognized until later (Georgi et al., 1990; Kingsley, 1994)). Importantly, DAF-4 was shown to bind human BMP ligands in cell culture, illustrating the conservation of these pathway components from nematodes to humans (Estevez et al., 1993). daf-4 is the sole type II receptor gene in C. elegans; it functions both in the DAF-7 pathway and in the DBL-1 pathway by interacting with different pathway-specific components (Estevez et al., 1993).
daf-4 is expressed widely during all stages of C. elegans development (Gunther et al., 2000; Patterson et al., 1997). Alternative polyadenylation of the daf-4 locus produces a truncated protein containing only the extracellular domain that acts as a negative regulator of DAF-7 function (Gunther and Riddle, 2004). daf-1, the type I receptor, is also widely expressed, primarily in the nervous system (Gunther et al., 2000). Expression of daf-1/type I receptor in interneurons is sufficient to rescue the dauer, egg-laying, feeding rate, and fat storage defects of a daf-1 mutant (Greer et al., 2008). Genetic evidence suggests that DAF-1/type I receptor is capable of signaling, albeit weakly, in the absence of phosphorylation by DAF-4/type II receptor. In a daf-4 mutant background, depletion of daf-1 causes a more severe mutant phenotype and overexpression of daf-1 partially rescues the daf-4 mutant phenotype (Gunther et al., 2000). Canonically, type I receptors differ from the constitutively active type II receptors in requiring phosphorylation for activation. The ability of DAF-1 to be constitutively active is supported by the phylogeny of TGF-β receptors that indicates that DAF-1 is somewhat related to both type I and type II receptors (Newfeld et al., 1999).
At the receiving cell, DAF-7 signaling is negatively regulated by BRA-1, which binds DAF-1 (Morita et al., 2001). BRA-1 was identified by homology to human BMP receptor-associated molecule BRAM1, which negatively regulates BMP signaling by binding BMP receptor-1A (Kurozumi et al., 1998). DAF-7 pathway receptor activity also may be modulated intracellularly by OBR-3 (originally published as BIP), an oxysterol-binding domain protein that binds BRA-1 and BRA-2/BMP receptor associated protein (BRAM) (Sugawara et al., 2001). However, knockdown of OBR-3 function results in small animals with no phenotypes associated with the DAF-7 pathway, suggesting a role for OBR-3 in DBL-1 signaling instead.
The Smads participating in this pathway are encoded by daf-8, daf-14, and daf-3. DAF-8 and DAF-14 are related to TGF-β R-Smads, except that DAF-14 lacks strong homology to the MH1 domain (Inoue and Thomas, 2000a; Park et al., 2010). These R-Smads have been shown to have partially redundant functions (Inoue and Thomas, 2000a; Park et al., 2010). Both of these Smads are expressed broadly. The Ser/Thr phosphatase PDP-1 has been identified as a negative regulator of DAF-8 and DAF-14 Smad activity (Narasimhan et al., 2011). In Drosophila and in mammals, PDP has similarly been identified as a Mad/Smad1 C-terminal phosphatase (Chen et al., 2006). C. elegans PDP-1 also modulates phenotypes of insulin signaling and nuclear localization of DAF-16/FoxO, providing evidence of crosstalk between the insulin and TGF-β pathways (see section 5.2) (Narasimhan et al., 2011).
daf-3 encodes a Smad distantly related to Co-Smads (Patterson et al., 1997). While DAF-8 and DAF-14 R-Smads function to transduce the DAF-7 signal, DAF-3 Co-Smad activity is negatively regulated by the pathway. In the absence of DAF-3, animals enter the non-dauer L3 stage regardless of the presence or absence of DAF-7 signaling (Patterson et al., 1997). Thus, while DAF-7 pathway signaling normally inhibits daf-3 activity it is dispensable in the absence of daf-3. Furthermore, DAF-3 drives a negative feedback loop by repressing the expression of daf-7 and daf-8 (Park et al., 2010). These antagonistic interactions differ from the mechanisms of inhibition by I-Smads in other systems.
daf-5 has the same type of genetic interactions with the DAF-7 pathway as daf-3. Cloning of daf-5 revealed homology to the Sno/Ski family of oncogenes that negatively regulate TGF-β signaling in vertebrates (da Graca et al., 2004; Tewari et al., 2004). Ski has been shown in vertebrates to be a component of the histone deacetylase complex (HDAC) and binds Smads (Akiyoshi et al., 1999; Luo et al., 1999; Sun et al., 1999; Wang et al., 2000; Xu et al., 2000). DAF-5 physically interacts with DAF-3 to promote dauer development, and may act in a conserved fashion as other Sno/Ski members (Tewari et al., 2004). Ski may repress TGF-β superfamily signaling in two ways. First, Ski recruits the HDAC to maintain the chromatin structure around TGF-β target genes in a transcriptionally silent form (Akiyoshi et al., 1999; Luo et al., 1999). Second, Ski prevents target gene transcription by stabilizing inactive Smad complexes on Smad-binding elements in the regulatory regions of target genes (Akiyoshi et al., 1999; Luo et al., 1999; Sun et al., 1999; Suzuki et al., 2004; Xu et al., 2000).
Microarray analyses have been performed to identify transcriptional target genes of the DAF-7 pathway. Liu et al. compared expression levels in daf-7, daf-8, and daf-14 mutant to wild-type populations during larval stages when the dauer decision was made and executed (Liu et al., 2004). Functional categories of identified target genes include insulin pathway genes, hedgehog- and patched-related genes, hormone biosynthetic genes, and cell cycle genes. Shaw et al. compared expression in adulthood in daf-7 and daf-1 mutant populations with expression in daf-3 and daf-5 Daf-d strains and in daf-7; daf-3 double mutant strains (Shaw et al., 2007). The adult targets of the pathway showed relatively low correlation with the dauer targets, and instead were enriched in functional categories that regulate longevity such as regulators of metabolism. Both studies identified a large degree of overlap with transcriptional targets of the insulin pathway at the same developmental stages. Consistent with this outcome, the upstream sequences of many of the target genes contained binding sites for DAF-16/FoxO, the transcription factor that mediates insulin signaling.
The DAF-7 pathway also regulates chemoreceptor gene expression in sensory neurons. Mutations in this pathway were identified in a screen for altered expression of the chemoreceptor gene sra-6/serpentine receptor, class A (Nolan et al., 2002). Several additional chemoreceptor genes normally expressed in the ASH and ASI sensory neurons or in the AWC sensory neurons also depend on DAF-7 pathway function for expression (Lesch and Bargmann, 2010; Nolan et al., 2002). Chemoreceptor gene regulation is likely mediated directly by DAF-3, since DAF-3 binding sites are prevalent upstream of chemoreceptor genes (McCarroll et al., 2005).
In large-scale interactome mapping analysis of this pathway using the yeast two-hybrid system, physical interactions among 59 proteins, including DAF-1/type I receptor, DAF-4/type II receptor, DAF-3/Smad, DAF-5/Sno/Ski, and several novel interactors were identified (Tewari et al., 2004). This work showed a physical interaction between DAF-3 and DAF-5. Among the novel interactors was DAF-21/HSP90, which interacts physically with DAF-1 and DAF-4 receptors. DAF-21 was previously shown to function in the DAF-11 cyclic nucleotide pathway, which processes dauer cues in parallel to the DAF-7 pathway. Thus, DAF-21 may act in multiple dauer-regulating pathways, possibly to ensure proper protein folding at the high temperatures that trigger dauer entry. TTX-1, a homeodomain protein required for thermosensation, is linked with both DAF-3 Smad and DAF-4 type II receptor indirectly via secondary interactions. DAF-3 also shows direct or indirect associations with SWI3 and SNF5 homologs (Y11G7B.23 and R07E5.3, respectively), suggesting a function in chromatin regulation.
In this section, we summarize the similarities between the DBL-1 and DAF-7 pathways as well as the differences that may reflect functional specializations. Both DBL-1 and DAF-7 ligands function alone, with no evidence of functional heterodimers. Both ligands interact with the sole type II receptor in C. elegans, DAF-4, with specificity determined by the type I receptor recruited (SMA-6 or DAF-1, respectively). In both pathways, two different R-Smads are required for full pathway activity, consistent with the functional Smad complex being a heterotrimer composed of three different subunits. However, while R-Smads SMA-2 and SMA-3 cannot replace one another, R-Smads DAF-8 and DAF-14 show some functional redundancy. In addition, each pathway has some biological roles in which a single R-Smad appears to function alone: e.g. SMA-3 in pathogen resistance and DAF-8 in germline mitosis. Unlike in vertebrates, C. elegans Co-Smads are pathway-specific. The Co-Smad SMA-4 is similar to mammalian Smad4 in both sequence and function. On the other hand, DAF-3 has sequence and functional features of both Co-Smads and I-Smads. In spite of the diversity of biological roles of these two pathways, the microarray analyses identified many of the same families of downstream target genes, with different individual genes represented in each case. These common families of target genes notably include those of the Hedgehog-related pathway and insulin-related ligand genes. Finally, both DBL-1 and DAF-7 pathways are controlled by tissue- and pathway-specific regulators (i.e., CRM-1, SMA-10, DRAG-1, LON-2 for DBL-1; BRA-1 and DAF-5 for DAF-7). As investigators continue to probe the functions of these pathways, it is likely that the number of modulators identified will continue to increase.
Unlike the other C. elegans TGF-β-related proteins, UNC-129 is a more ancestral protein with no obvious subfamily affiliation based on sequence or in vivo function (Colavita et al., 1998). Also unique to UNC-129 is that it has no recognized type I and type II receptors. Rather, as confirmed in tissue culture, UNC-129 interacts with the UNC-5 netrin receptor, a transmembrane protein with two immunoglobulin-like domains and two type I thrombospondin motifs in the extracellular region that shares no obvious similarity with other TGF-β receptors (MacNeil et al., 2009).
UNC-129 is expressed in several tissues, including dorsal body wall muscle, where it facilitates the long-range migrations of motor neurons and distal tip cells (DTCs) (Colavita et al., 1998; Gabel et al., 2008; MacNeil et al., 2009). Too little or too much UNC-129 causes misguided PVPR and PVQL, which pioneer the left axon tract, and motor axons (DA, DB, VD, DD, NSM, and SDQ), producing the namesake uncoordinated (Unc) phenotype (Axäng et al., 2008; Colavita and Culotti, 1998; Colavita et al., 1998; Hedgecock et al., 1990; Qin and Powell-Coffman, 2004). UNC-129 also plays a minor role in the migration of the pharyngeal M2 motor neuron (Mörck et al., 2003). This role is facilitated by MNM-2 (M neuron morphology abnormal-2), a Krüppel-like zinc finger protein that affects the M2 sister cell M3, which helps guide M2 (Rauthan et al., 2007). Although UNC-129 is required for neuronal guidance during development, it does not appear to be required for axon guidance during regeneration (Gabel et al., 2008).
The two DTCs also use UNC-129 in their U-shaped migrations that pattern the hermaphrodite gonad (Merz et al., 2003). UNC-129 in the dorsal body wall muscle attracts the DTCs, promoting their lateral turn from the ventral muscle band and migration toward the dorsal muscle band (distal tip cell migration phase 2) (Merz et al., 2003). Although loss of UNC-129 does not by itself affect DTC migrations, the need for UNC-129 is revealed when levels of its receptor, UNC-5, are also reduced (MacNeil et al., 2009; Merz et al., 2003). Some gonads of animals with hypomorphic unc-5 and loss of unc-129 expression have both proximal and distal gonad ends positioned on the dorsal side, instead of having only the proximal end dorsally positioned and the distal end ventrally positioned. Ectopic expression of UNC-129 in ventral body wall muscle instead of its normal dorsal body wall muscle site also causes DTCs to mis-migrate (Colavita et al., 1998; Kinnunen et al., 2005; Nash et al., 2000). An unc-129 null mutation partially suppresses the enhancement by class I unc-52/perlecan alleles of unc-5(e152) DTC migration defects, suggesting that UNC-52, an extracellular matrix heparan sulfate proteoglycan, acts in part through UNC-129 to direct DTC migration (Merz et al., 2003). This observation is supported by findings in other systems that perlecan modulates TGF-β bio-availability and activity (Lamoureux et al., 2007).
In addition to its axon guidance role, a role for unc-129 in male tail sensory ray development has been identified (Ikegami et al., 2004). UNC-129 is expressed in specific ray neurons (rays 1, 3, 5, and 7) and their precursor Rn cells (Ikegami et al., 2004; Nash et al., 2000). Alteration of UNC-129 levels alone has no effect on rays or the formation of ray axon commissures, axon processes that run from one side of the body to the other circumferentially (Ikegami et al., 2004; Jia and Emmons, 2006; Roy et al., 2000). However, a role for UNC-129 is revealed in animals lacking gene products required for cell separation and neuron migrations. unc-129 interacts genetically with the semaphorin 2a gene mab-20 and the semaphorin receptor gene plx-2/plexin, and acts in parallel with efn-4/ephrin, to prevent sensory ray fusions between adjacent rays.
UNC-129 attracts cells by promoting the formation of UNC-5/UNC-40 receptor heterodimers, which promote long-range cell migration, at the expense of UNC-5 homodimers, which promote short-range migration (Colavita et al., 1998; Gabel et al., 2008; MacNeil et al., 2009). Expression of unc-129 in ventral body muscles is normally inhibited by the forkhead transcription factor UNC-130 (Nash et al., 2000). Loss of UNC-130 leads to a phenotype that closely resembles that of animals ectopically overexpressing unc-129 in both dorsal and ventral body wall muscles (Nash et al., 2000). unc-129 expression may also require the T-box transcription factor MAB-9, as loss of mab-9 reduces expression of a short unc-129 promoter fusion with GFP that expresses in DA and SB motor neurons in transgenic animals (Colavita et al., 1998; Pocock et al., 2008). The extracellular matrix constituent TEN-1/teneurin, a single-pass transmembrane protein with EGF-like domains, also appears to normally restrict the expression of unc-129 in the dorsal muscle, as an unc-129 gene fusion with GFP is 20-fold upregulated in ten-1 loss-of-function animals (Mörck et al., 2010).
In other systems, many TGF-β superfamily members act redundantly. In C. elegans, there is some overlap in function between DBL-1 and DAF-7. DBL-1 and DAF-7 pathways appear to both work to maintain a non-dauered animal. Expression of both is lost upon entering dauer conditions. Dauering by loss of DAF-7 pathway function reduces dbl-1 expression (Liu et al., 2004), and daf-7 is normally repressed in dauering conditions (Schackwitz et al., 1996). Not only does the DBL-1 pathway respond to dauering cues, but it appears to play a small role in preventing dauering. The daf-1 Daf-c phenotype is enhanced in double mutant strains sma-6(lf); daf-1(lf) (Krishna et al., 1999; Maduzia et al., 2005) and daf-7(lf); dbl-1(lf) (Morita et al., 1999), indicating that sma-6 and dbl-1 normally function at a low level to repress dauer-promoting signaling.
DBL-1 and UNC-129 pathways both promote proper distal tip cell (DTC) migratory paths through UNC-5/netrin receptor (Merz et al., 2003; Nash et al., 2000). DBL-1, with receptor DAF-4, is proposed to act on the ventral side as a DTC repellant, while UNC-129 acts on the dorsal side to attract DTCs (Nash et al., 2000). This guidance role is not seen in animals where the function of either of these TGF-βs is lost. However, in a hypomorphic unc-5 mutant background, mutation of either dbl-1 or unc-129 causes more DTC slipping than the unc-5 hypomorphic background alone (Merz et al., 2003).
TGF-β pathways interact with other pathways to modulate signaling output, including insulins and Hedgehog-related warthogs (Guo and Wang, 2009). C. elegans provides a useful model system to study TGF-β signaling pathways relatively uncomplicated by pathways that can confound analyses in other systems. Many signaling pathways have fewer members in C. elegans than in other metazoans and provide clear, distinguishable phenotypes (Pires-daSilva and Sommer, 2003). For example, C. elegans contains highly diverged Hedgehog pathway members, which interact with TGF-β at the level of the receptors and/or Smads in other systems, but have no defined role in TGF-β signaling in the nematode (Hao et al., 2006). Largely non-overlapping Hedgehog-related pathways appear to be transcriptionally regulated by DBL-1 or DAF-7 signaling and may be players in transducing TGF-β superfamily signals (Liang et al., 2007; Liu et al., 2004; Roberts et al., 2010). The Hedgehog-related factors upregulated by DBL-1 signaling are required for normal body size (Roberts et al., 2010; Zugasti et al., 2005). Other signaling pathways, including Wnt/Wg, EGF, and FGF, exist in the nematode but act in tissues distinct from those associated with TGF-β signaling (see Wnt Signaling; Genomic overview of protein kinases; Vulval development). In this section, we review some of the molecular pathway interactions that are conserved in other systems.
Insulin pathway interactions
In genetic studies of dauer development, three interacting signaling pathways were found to be involved: the DAF-7 pathway, an insulin signaling pathway, and a cyclic nucleotide pathway. In addition to their shared role in dauer development (see section 3.1), the insulin and DAF-7 pathways also possess common functions in the regulation of fat metabolism and longevity (see sections 3.2 and 3.4). In regulation of dauer development, inputs from the DAF-7 and insulin pathways must be integrated to determine the organism's response. Some of this integration is mediated by the nuclear hormone receptor DAF-12 (Antebi et al., 2000; Snow and Larsen, 2000). Like daf-3 and daf-5, daf-12 mutations suppress the DAF-7 pathway Daf-c phenotype. Unlike daf-3 and daf-5, however, daf-12 also shows strong suppression of mutations in the insulin-related and cyclic nucleotide pathways and is required for regulating multiple stage-specific aspects of nematode development (Antebi et al., 2000; Snow and Larsen, 2000). The daf-12 locus is complex, encoding multiple alternatively spliced isoforms, and is capable of mutating to Daf-d, Daf-c, and heterochronic phenotypes independently. This structural complexity may contribute to its functional complexity. Crosstalk between the TGF-β and insulin dauer pathways may also be mediated in part by the regulation of insulin ligand expression by the DAF-7 pathway (Liu et al., 2004; Shaw et al., 2007). Finally, the DAF-7 pathway promotes secretion of the insulin-like ligand DAF-28 (Park et al., 2012a). This effect is mediated by an interaction between DAF-8/Smad and NHR-69/nuclear hormone receptor that represses expression of exp-2, a potassium channel gene, to promote DAF-28 secretion. Thus, DAF-7/TGF-β signaling can promote insulin pathway activity via multiple mechanisms.
Interaction of the insulin pathway with the DBL-1 pathway has been less well studied. The DBL-1 and insulin pathways both regulate reproductive aging, but they appear to do so in parallel (Luo et al., 2010). On the other hand, the DBL-1 pathway, like the DAF-7 pathway, regulates the expression of insulin-like ligands (Liang et al., 2007; Luo et al., 2010). Furthermore, the DBL-1 pathway regulates the expression of a subset of known DAF-16/FOXO transcriptional targets such as fat-6 and fat-7 (Liang et al., 2007). Finally, some daf-2/insulin receptor mutant animals have increased body size, suggesting an antagonistic interaction with the DBL-1 pathway in the regulation of growth (McCulloch and Gems, 2003). Additional studies will be necessary to determine whether these observations reflect a relevant regulatory interaction between the DBL-1 and insulin pathways.
Chemosensory pathways
Some signaling pathways and physiological processes affect body size by means that may partially merge with DBL-1 signaling. For example, several genes that function in chemosensation result in alterations in body size. These include tax-6 and cnb-1, which encode the catalytic and regulatory subunits of calcineurin, respectively. Calcineurin is a protein phosphatase 2B (PP2B) that controls intracellular Ca2+ signaling and mediates transcriptional responses responsible for a variety of physiological processes in many systems. tax-6 interacts with kin-29 (a.k.a. sma-11), which encodes a serine/threonine kinase that is a downstream target of DBL-1 signaling (see section 2.8) (Maduzia et al., 2005; Singaravelu et al., 2007).
egl-4, a gene encoding cGMP-dependent protein kinase, acts downstream of chemosensory genes to regulate body size by repressing the DBL-1 pathway (Fujiwara et al., 2002; Hirose et al., 2003). egl-4 is expressed in neurons, hypodermis, intestine, and body wall muscles (Fujiwara et al., 2002; Hirose et al., 2003; L'Etoile et al., 2002). Expression of egl-4 in neurons or in hypodermis can rescue the organ size defects nonautonomously (Nakano et al., 2004). Interestingly, EGL-4 protein directly interacts with KIN-29 (Lanjuin and Sengupta, 2002; Maduzia et al., 2005; van der Linden et al., 2008). egl-4 also acts downstream of chemosensory genes to regulate dauer entry and other metabolic responses (see sections 3.2 and 3.3).
Receptor tyrosine kinase signaling
scd-2 (suppressor of constitutive dauer-2) mutant alleles were identified in a genetic screen for suppressors of the TGF-β dauer-constitutive phenotype (Inoue and Thomas, 2000b) and as wild isolates from an oasis in a California desert (Reiner et al., 2008). SCD-2 is the C. elegans ortholog of anaplastic lymphoma kinase (ALK), a receptor tyrosine kinase. SCD-2, its ligand HEN-1, the adaptor SOC-1, and the downstream MAPK SMA-5 regulate dauer development upstream of DAF-3/Co-Smad but in parallel with DAF-7 (Reiner et al., 2008). The SCD-2 pathway is specific to the DAF-3-mediated dauer response, as it has no effect on the other phenotypes of the DAF-7 pathway.
In addition to MAPK pathways, receptor tyrosine kinases can also signal through the STAT pathway (Levy and Darnell, 2002). The C. elegans STAT homolog STA-1 is expressed in chemosensory neurons in the head, and its expression is negatively regulated by daf-7, daf-4, and daf-8 (Wang and Levy, 2006). Furthermore, loss of sta-1 activity enhances the dauer-constitutive phenotypes of daf-7, daf-4, and daf-14 mutant animals, but not of daf-1 and daf-8 mutants. These results indicate that STA-1 negatively regulates dauer formation in a DAF-1- and DAF-8-dependent manner, and points to the existence of crosstalk between the STAT and TGF-β pathways.
In humans, misregulation of TGF-β superfamily pathways leads to developmental defects and diseases, such as vascular defects, bone abnormalities including dwarfism, overgrowth syndromes including Marfan syndrome, midline defects, and cancers (Padua and Massagué, 2009; Serra and Chang, 2003; Wu and Hill, 2009). What challenges remain for the future that can be addressed using the C. elegans model system? We can anticipate that new genetic, molecular, and chemical screens will uncover additional protein, RNA, and lipid molecules that modulate TGF-β signaling, as well as new functions for known signaling components. New factors may function to stimulate or restrict TGF-β signaling in the sending cell, in the extracellular space, or in the cytoplasm or the nucleus of the receiving cell. Furthermore, studies of the transcriptional outputs of TGF-β signaling in C. elegans are still in their infancy, while studies of non-transcriptional outputs have, for the most part, not yet been carried out. These areas of research will shed light on the spectrum of possible cellular responses to TGF-β signaling, identifying both conserved and nematode-specific physiological roles.
An advantage of working in a genetic model system such as C. elegans is the ability to test perturbations within the context of the intact animal. For this reason, we expect that research in C. elegans will continue to provide insight into the difficult questions of how TGF-β signaling is integrated with other signaling pathways, with developmental and cell cycle timing, and with environmental cues. Furthermore, a critical area of study for the TGF-β field is to understand how TGF-β signaling achieves such remarkable context dependence. For example, many TGF-β superfamily ligands act as morphogens during development, specifying different outcomes at different concentrations (McDowell and Gurdon, 1999; Podos and Ferguson, 1999). In addition, TGF-β itself plays dual, opposing roles in cancer progression: as a tumor suppressor early in tumor progression and then as a pro-oncogenic factor later in carcinogenesis (Roberts and Wakefield, 2003). By studying how two simple TGF-β signaling pathways can achieve a wide variety of biological outcomes in C. elegans, mechanisms for context dependence will be identified that can then be tested in other systems. In summary, the simple, yet powerful, model system C. elegans promises to continue to provide insight into evolutionarily conserved molecules and mechanisms relevant to understanding not only dynamic cellular processes but also pathological states including developmental disorders and cancers.
Work in the Gumienny laboratory is supported in part by Beginning Grant-in-Aid 11BGIA5700014 from the American Heart Association – South Central Affiliate and NIH 1R01GM097591-01. Work in the Savage-Dunn laboratory is supported in part by 1R15GM073678 and 1R15GM097692 from NIH. We thank Suparna Bageshwar, Robbie Schultz, Kathy Beifuss, Jun Liang Rice, Daniel Weinstein, Robyn Lints, Leon Avery, and Jane Hubbard for discussions and critical reading of the manuscript.
Ailion, M., and Thomas, J.H. (2000). Dauer formation induced by high temperatures in Caenorhabditis elegans. Genetics 156, 1047-1067. Abstract
Akiyoshi, S., Inoue, H., Hanai, J., Kusanagi, K., Nemoto, N., Miyazono, K., and Kawabata, M. (1999). c-Ski acts as a transcriptional co-repressor in transforming growth factor-β signaling through interaction with smads. J. Biol. Chem. 274, 35269-35277. Abstract Article
Almedom, R.B., Liewald, J.F., Hernando, G., Schultheis, C., Rayes, D., Pan, J., Schedletzky, T., Hutter, H., Bouzat, C., and Gottschalk, A. (2009). An ER-resident membrane protein complex regulates nicotinic acetylcholine receptor subunit composition at the synapse. EMBO J. 28, 2636-2649. Abstract Article
Alper, S., McBride, S.J., Lackford, B., Freedman, J.H., and Schwartz, D.A. (2007). Specificity and complexity of the Caenorhabditis elegans innate immune response. Mol. Cell. Biol. 27, 5544-5553. Abstract Article
Andriopoulos, B., Jr., Corradini, E., Xia, Y., Faasse, S.A., Chen, S., Grgurevic, L., Knutson, M.D., Pietrangelo, A., Vukicevic, S., Lin, H.Y., et al. (2009). BMP6 is a key endogenous regulator of hepcidin expression and iron metabolism. Nat. Genet. 41, 482-487. Abstract Article
Antebi, A., Yeh, W.H., Tait, D., Hedgecock, E.M., and Riddle, D.L. (2000). daf-12 encodes a nuclear receptor that regulates the dauer diapause and developmental age in C. elegans. Genes Dev. 14, 1512-1527. Abstract Article
Ashrafi, K. Obesity and the regulation of fat metabolism (March 9, 2007), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.130.1, http://www.wormbook.org. Article
Austin, J., and Kimble, J. (1987). glp-1 is required in the germ line for regulation of the decision between mitosis and meiosis in C. elegans. Cell, 589-599. Article
Axäng, C., Rauthan, M., Hall, D.H., and Pilon, M. (2008). Developmental genetics of the C. elegans pharyngeal neurons NSML and NSMR. BMC Dev. Biol. 8, 38. Abstract Article
Babitt, J.L., Huang, F.W., Xia, Y., Sidis, Y., Andrews, N.C., and Lin, H.Y. (2007). Modulation of bone morphogenetic protein signaling in vivo regulates systemic iron balance. J. Clin. Invest. 117, 1933-1939. Abstract Article
Baird, S.E., and Ellazar, S.A. (1999). TGFβ-like signaling and spicule development in Caenorhabditis elegans. Dev. Biol. 212, 93-100. Abstract Article
Baldessari, D., Badaloni, A., Longhi, R., Zappavigna, V., and Consalez, G.G. (2004). MAB21L2, a vertebrate member of the Male-abnormal 21 family, modulates BMP signaling and interacts with SMAD1. BMC Cell Biol. 5, 48. Abstract Article
Bargmann, C.I. Chemosensation in C. elegans (October 25, 2006), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.123.1, http://www.wormbook.org. Article
Ben Arous, J., Laffont, S., and Chatenay, D. (2009). Molecular and sensory basis of a food related two-state behavior in C. elegans. PLoS One 4, e7584. Abstract Article
Brand, A.M., Varghese, G., Majewski, W., and Hawdon, J.M. (2005). Identification of a DAF-7 ortholog from the hookworm Ancylostoma caninum. Int. J. Parasitol. 35, 1489-1498. Abstract Article
Brenner, S. (1974). The genetics of Caenorhabditis elegans. Genetics 77, 71-94. Abstract
Cai, Q., Wang, W., Gao, Y., Yang, Y., Zhu, Z., and Fan, Q. (2009). Ce-wts-1 plays important roles in Caenorhabditis elegans development. FEBS Lett. 583, 3158-3164. Abstract Article
Cassada, R.C., and Russell, R.L. (1975). The dauerlarva, a post-embryonic developmental variant of the nematode Caenorhabditis elegans. Dev. Biol. 46, 326-342. Abstract Article
Chang, A.J., Chronis, N., Karow, D.S., Marletta, M.A., and Bargmann, C.I. (2006). A distributed chemosensory circuit for oxygen preference in C. elegans. PLoS Biol. 4. Abstract Article
Chase, D.L. and Koelle, M.R. Biogenic amine neurotransmitters in C. elegans (February 20, 2007), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.132.1, http://www.wormbook.org. Article
Chen, H.B., Shen, J., Ip, Y.T., and Xu, L. (2006). Identification of phosphatases for Smad in the BMP/DPP pathway. Genes Dev. 20, 648-653. Abstract Article
Chow, K.L., Hall, D.H., and Emmons, S.W. (1995). The mab-21 gene of Caenorhabditis elegans encodes a novel protein required for choice of alternate cell fates. Development 121, 3615-3626. Abstract
Choy, S.W., Wong, Y.M., Ho, S.H., and Chow, K.L. (2007). C. elegans SIN-3 and its associated HDAC corepressor complex act as mediators of male sensory ray development. Biochem. Biophys. Res. Commun. 358, 802-807. Abstract Article
Colavita, A., and Culotti, J.G. (1998). Suppressors of ectopic UNC-5 growth cone steering identify eight genes involved in axon guidance in Caenorhabditis elegans. Dev. Biol. 194, 72-85. Abstract Article
Colavita, A., Krishna, S., Zheng, H., Padgett, R.W., and Culotti, J.G. (1998). Pioneer axon guidance by UNC-129, a C. elegans TGF-β. Science 281, 706-709. Abstract Article
Colige, A., Beschin, A., Samyn, B., Goebels, Y., Van Beeumen, J., Nusgens, B.V., and Lapiere, C.M. (1995). Characterization and partial amino acid sequencing of a 107-kDa procollagen I N-proteinase purified by affinity chromatography on immobilized type XIV collagen. J. Biol. Chem. 270, 16724-16730. Abstract Article
Collins, J.J., et al. The measurement and analysis of age-related changes in Caenorhabditis elegans (December 7, 2007), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.137.1, http://www.wormbook.org. Article
Collins-Racie, L.A., Flannery, C.R., Zeng, W., Corcoran, C., Annis-Freeman, B., Agostino, M.J., Arai, M., DiBlasio-Smith, E., Dorner, A.J., Georgiadis, K.E., et al. (2004). ADAMTS-8 exhibits aggrecanase activity and is expressed in human articular cartilage. Matrix Biol. 23, 219-230. Abstract Article
Corradini, E., Babitt, J.L., and Lin, H.Y. (2009). The RGM/DRAGON family of BMP co-receptors. Cytokine Growth Factor Rev. 20, 389-398. Abstract Article
Crook, M., Grant, K., and Grant, W.N. (2010). Failure of Parastrongyloides trichosuri daf-7 to complement a Caenorhabditis elegans daf-7 (e1372) mutant: implications for the evolution of parasitism. Int. J. Parasitol. 40, 1675-1683. Abstract Article
Crook, M., Thompson, F.J., Grant, W.N., and Viney, M.E. (2005). daf-7 and the development of Strongyloides ratti and Parastrongyloides trichosuri. Mol. Biochem. Parasitol. 139, 213-223. Abstract Article
da Graca, L.S., Zimmerman, K.K., Mitchell, M.C., Kozhan-Gorodetska, M., Sekiewicz, K., Morales, Y., and Patterson, G.I. (2004). DAF-5 is a Ski oncoprotein homolog that functions in a neuronal TGFβ pathway to regulate C. elegans dauer development. Development 131, 435-446. Abstract Article
Dalfó, D., Michaelson, D., and Hubbard, E.J. (2012). Sensory regulation of the C. elegans germline through TGF-β-dependent signaling in the niche. Curr. Biol., 712-719. Abstract Article
Daniels, S.A., Ailion, M., Thomas, J.H., and Sengupta, P. (2000). egl-4 acts through a transforming growth factor-β/SMAD pathway in Caenorhabditis elegans to regulate multiple neuronal circuits in response to sensory cues. Genetics 156, 123-141. Abstract
de Bono, M., and Bargmann, C.I. (1998). Natural variation in a neuropeptide Y receptor homolog modifies social behavior and food response in C. elegans. Cell 94, 679-689. Abstract Article
De Robertis, E.M. (2008). Evo-devo: variations on ancestral themes. Cell 132, 185-195. Abstract Article
Duerr, J.S., Han, H.P., Fields, S.D., and Rand, J.B. (2008). Identification of major classes of cholinergic neurons in the nematode Caenorhabditis elegans. J. Comp. Neurol. 506, 398-408. Abstract Article
Eisenmann, D. M., Wnt signaling (June 25, 2005), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.7.1, http://www.wormbook.org. Article
Emmons, S.W. Male development (November 10, 2005), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.33.1, http://www.wormbook.org. Article
Estevez, M., Attisano, L., Wrana, J.L., Albert, P.S., Massagué, J., and Riddle, D.L. (1993). The daf-4 gene encodes a bone morphogenetic protein receptor controlling C. elegans dauer larva development. Nature 365, 644-649. Abstract Article
Estevez, M., Estevez, A.O., Cowie, R.H., and Gardner, K.L. (2004). The voltage-gated calcium channel UNC-2 is involved in stress-mediated regulation of tryptophan hydroxylase. J. Neurochem. 88, 102-113. Abstract Article
Ewbank, J. J. Signaling in the immune response (January 23, 2006), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.83.1, http://www.wormbook.org. Article
Fernandes, R.J., Hirohata, S., Engle, J.M., Colige, A., Cohn, D.H., Eyre, D.R., and Apte, S.S. (2001). Procollagen II amino propeptide processing by ADAMTS-3. Insights on dermatosparaxis. J. Biol. Chem. 276, 31502-31509. Abstract Article
Fernando, T., Flibotte, S., Xiong, S., Yin, J., Yzeiraj, E., Moerman, D.G., Meléndez, A., and Savage-Dunn, C. (2011). C. elegans ADAMTS ADT-2 regulates body size by modulating TGFβ signaling and cuticle collagen organization. Dev. Biol. 352, 92-103. Abstract Article
Flemming, A.J., Shen, Z.Z., Cunha, A., Emmons, S.W., and Leroi, A.M. (2000). Somatic polyploidization and cellular proliferation drive body size evolution in nematodes. Proc. Natl. Acad. Sci. U. S. A. 97, 5285-5290. Abstract Article
Foehr, M.L., Lindy, A.S., Fairbank, R.C., Amin, N.M., Xu, M., Yanowitz, J., Fire, A.Z., and Liu, J. (2006). An antagonistic role for the C. elegans Schnurri homolog SMA-9 in modulating TGFβ signaling during mesodermal patterning. Development 133, 2887-2896. Abstract Article
Foehr, M.L., and Liu, J. (2008). Dorsoventral patterning of the C. elegans postembryonic mesoderm requires both LIN-12/Notch and TGFβ signaling. Dev. Biol. 313, 256-266. Abstract Article
Freitas, T.C., and Arasu, P. (2005). Cloning and characterisation of genes encoding two transforming growth factor-β-like ligands from the hookworm, Ancylostoma caninum. Int. J. Parasitol. 35, 1477-1487. Abstract Article
Fujiwara, M., Sengupta, P., and McIntire, S.L. (2002). Regulation of body size and behavioral state of C. elegans by sensory perception and the EGL-4 cGMP-dependent protein kinase. Neuron 36, 1091-1102. Abstract Article
Fung, W.Y., Fat, K.F., Eng, C.K., and Lau, C.K. (2007). crm-1 facilitates BMP signaling to control body size in Caenorhabditis elegans. Dev. Biol. 311, 95-105. Abstract Article
Gabel, C.V., Antoine, F., Chuang, C.F., Samuel, A.D., and Chang, C. (2008). Distinct cellular and molecular mechanisms mediate initial axon development and adult-stage axon regeneration in C. elegans. Development 135, 1129-1136. Abstract Article
Georgi, L.L., Albert, P.S., and Riddle, D.L. (1990). daf-1, a C. elegans gene controlling dauer larva development, encodes a novel receptor protein kinase. Cell 61, 635-645. Abstract Article
Golden, J.W., and Riddle, D.L. (1984a). The Caenorhabditis elegans dauer larva: developmental effects of pheromone, food, and temperature. Dev. Biol. 102, 368-378. Abstract Article
Golden, J.W., and Riddle, D.L. (1984b). A pheromone-induced developmental switch in Caenorhabditis elegans: Temperature-sensitive mutants reveal a wild-type temperature-dependent process. Proc. Natl. Acad. Sci. U. S. A. 81, 819-823. Abstract Article
Golden, T.R. and Melov, S. Gene expression changes associated with aging in C. elegans (February 12, 2007), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.127.2, http://www.wormbook.org. Article
Goldsmith, A.D., Sarin, S., Lockery, S., and Hobert, O. (2010). Developmental control of lateralized neuron size in the nematode Caenorhabditis elegans. Neural Dev. 5, 33. Abstract Article
Gomez-Escobar, N., Gregory, W.F., and Maizels, R.M. (2000). Identification of tgh-2, a filarial nematode homolog of Caenorhabditis elegans daf-7 and human transforming growth factor β, expressed in microfilarial and adult stages of Brugia malayi. Infect. Immun. 68, 6402-6410. Abstract Article
Gracheva, E.O., Burdina, A.O., Holgado, A.M., Berthelot-Grosjean, M., Ackley, B.D., Hadwiger, G., Nonet, M.L., Weimer, R.M., and Richmond, J.E. (2006). Tomosyn inhibits synaptic vesicle priming in Caenorhabditis elegans. PLoS Biol. 4, e261. Abstract Article
Gray, J.M., Karow, D.S., Lu, H., Chang, A.J., Chang, J.S., Ellis, R.E., Marletta, M.A., and Bargmann, C.I. (2004). Oxygen sensation and social feeding mediated by a C. elegans guanylate cyclase homologue. Nature 430, 317-322. Abstract Article
Greer, E.R., Perez, C.L., Van Gilst, M.R., Lee, B.H., and Ashrafi, K. (2008). Neural and molecular dissection of a C. elegans sensory circuit that regulates fat and feeding. Cell Metab. 8, 118-131. Abstract Article
Gumienny, T.L., Macneil, L., Zimmerman, C.M., Wang, H., Chin, L., Wrana, J.L., and Padgett, R.W. (2010). Caenorhabditis elegans SMA-10/LRIG is a conserved transmembrane protein that enhances bone morphogenetic protein signaling. PLoS Genet. 6, e1000963. Abstract Article
Gumienny, T.L., MacNeil, L.T., Wang, H., de Bono, M., Wrana, J.L., and Padgett, R.W. (2007). Glypican LON-2 is a conserved negative regulator of BMP-like signaling in Caenorhabditis elegans. Curr. Biol. 17, 159-164. Abstract Article
Gunther, C.V., Georgi, L.L., and Riddle, D.L. (2000). A Caenorhabditis elegans type I TGFβ receptor can function in the absence of type II kinase to promote larval development. Development 127, 3337-3347. Abstract
Gunther, C.V., and Riddle, D.L. (2004). Alternative polyadenylation results in a truncated daf-4 BMP receptor that antagonizes DAF-7-mediated development in Caenorhabditis elegans. J. Biol. Chem. 279, 39555-39564. Abstract Article
Guo, X., and Wang, X.F. (2009). Signaling cross-talk between TGF-β/BMP and other pathways. Cell Res. 19, 71-88. Abstract Article
Hahn, A.C., and Emmons, S.W. (2003). The roles of an ephrin and a semaphorin in patterning cell-cell contacts in C. elegans sensory organ development. Dev. Biol. 256, 379-388. Abstract Article
Halder, G., and Johnson, R.L. (2010). Hippo signaling: growth control and beyond. Development 138, 9-22. Abstract Article
Hanai, J., Chen, L.F., Kanno, T., Ohtani-Fujita, N., Kim, W.Y., Guo, W.H., Imamura, T., Ishidou, Y., Fukuchi, M., Shi, M.J., et al. (1999). Interaction and functional cooperation of PEBP2/CBF with Smads. Synergistic induction of the immunoglobulin germline Cα promoter. J. Biol. Chem. 274, 31577-31582. Abstract Article
Hao, L., Johnsen, R., Lauter, G., Baillie, D., and Burglin, T.R. (2006). Comprehensive analysis of gene expression patterns of hedgehog-related genes. BMC Genomics 7, 280. Abstract Article
Hedgecock, E.M., Culotti, J.G., and Hall, D.H. (1990). The unc-5, unc-6, and unc-40 genes guide circumferential migrations of pioneer axons and mesodermal cells on the epidermis in C. elegans. Neuron 4, 61-85. Abstract Article
Herpin, A., Lelong, C., and Favrel, P. (2004). Transforming growth factor-β-related proteins: an ancestral and widespread superfamily of cytokines in metazoans. Dev. Comp. Immunol. 28, 461-485. Abstract Article
Hirose, T., Nakano, Y., Nagamatsu, Y., Misumi, T., Ohta, H., and Ohshima, Y. (2003). Cyclic GMP-dependent protein kinase EGL-4 controls body size and lifespan in C elegans. Development 130, 1089-1099. Abstract Article
Ho, S.H., So, G.M., and Chow, K.L. (2001). Postembryonic expression of Caenorhabditis elegans mab-21 and its requirement in sensory ray differentiation. Dev. Dyn. 221, 422-430. Abstract Article
Horvitz, H.R., Chalfie, M., Trent, C., Sulston, J.E., and Evans, P.D. (1982). Serotonin and octopamine in the nematode Caenorhabditis elegans. Science 216, 1012-1014. Abstract Article
Hu, P.J. Dauer (August 08, 2007), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.144.1, http://www.wormbook.org. Article
Ikegami, R., Zheng, H., Ong, S.H., and Culotti, J. (2004). Integration of semaphorin-2A/MAB-20, ephrin-4, and UNC-129 TGF-β signaling pathways regulates sorting of distinct sensory rays in C. elegans. Dev. Cell 6, 383-395. Abstract Article
Immergluck, K., Lawrence, P.A., and Bienz, M. (1990). Induction across germ layers in Drosophila mediated by a genetic cascade. Cell 62, 261-268. Abstract Article
Inman, G.J., Nicolás, F.J., and Hill, C.S. (2002). Nucleocytoplasmic shuttling of Smads 2, 3, and 4 permits sensing of TGF-β receptor activity. Mol. Cell 10, 283-294. Abstract Article
Inoue, T., and Thomas, J.H. (2000a). Targets of TGF-β signaling in Caenorhabditis elegans dauer formation. Dev. Biol. 217, 192-204. Abstract Article
Inoue, T., and Thomas, J.H. (2000b). Suppressors of transforming growth factor-β pathway mutants in the Caenorhabditis elegans dauer formation pathway. Genetics 156, 1035-1046. Abstract
Inoue, T., Ailion, M., Poon, S., Kim, H.K., Thomas, J.H., and Sternberg, P.W. (2007). Genetic analysis of dauer formation in Caenorhabditis briggsae. Genetics 177, 809-818. Abstract Article
Inoue, Y., and Imamura, T. (2008). Regulation of TGF-β family signaling by E3 ubiquitin ligases. Cancer Sci. 99, 2107-2112. Abstract Article
Irving, P., Troxler, L., Heuer, T.S., Belvin, M., Kopczynski, C., Reichhart, J.M., Hoffmann, J.A., and Hetru, C. (2001). A genome-wide analysis of immune responses in Drosophila. Proc. Natl. Acad. Sci. U. S. A. 98, 15119-15124. Abstract Article
Itoh, S., Itoh, F., Goumans, M.J., and Ten Dijke, P. (2000). Signaling of transforming growth factor-β family members through Smad proteins. Eur. J. Biochem. 267, 6954-6967. Abstract Article
Ji, Y.J., Nam, S., Jin, Y.H., Cha, E.J., Lee, K.S., Choi, K.Y., Song, H.O., Lee, J., Bae, S.C., and Ahnn, J. (2004). RNT-1, the C. elegans homologue of mammalian RUNX transcription factors, regulates body size and male tail development. Dev. Biol. 274, 402-412. Abstract Article
Jia, L., and Emmons, S.W. (2006). Genes that control ray sensory neuron axon development in the Caenorhabditis elegans male. Genetics 173, 1241-1258. Abstract Article
Jones, K.T., Greer, E.R., Pearce, D., and Ashrafi, K. (2009). Rictor/TORC2 regulates Caenorhabditis elegans fat storage, body size, and development through sgk-1. PLoS Biol. 7, e60. Abstract Article
Jorgensen, E.M. GABA (August 31, 2005), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.14.1, http://www.wormbook.org. Article
Kenyon, C., Chang, J., Gensch, E., Rudner, A., and Tabtiang, R. (1993). A C. elegans mutant that lives twice as long as wild type. Nature 366, 461-464. Abstract Article
Kingsley, D.M. (1994). The TGF-β superfamily: new members, new receptors, and new genetic tests of function in different organisms. Genes Dev. 8, 133-146. Abstract Article
Kinnunen, T., Huang, Z., Townsend, J., Gatdula, M.M., Brown, J.R., Esko, J.D., and Turnbull, J.E. (2005). Heparan 2-O-sulfotransferase, hst-2, is essential for normal cell migration in Caenorhabditis elegans. Proc. Natl. Acad. Sci. U. S. A. 102, 1507-1512. Abstract Article
Kirkpatrick, C.A., Knox, S.M., Staatz, W.D., Fox, B., Lercher, D.M., and Selleck, S.B. (2006). The function of a Drosophila glypican does not depend entirely on heparan sulfate modification. Dev. Biol. 300, 570-582. Abstract Article
Kratsios, P., Stolfi, A., Levine, M., and Hobert, O. (2011). Coordinated regulation of cholinergic motor neuron traits through a conserved terminal selector gene. Nat. Neurosci. 15, 205-214. Abstract Article
Krishna, S., Maduzia, L.L., and Padgett, R.W. (1999). Specificity of TGFβ signaling is conferred by distinct type I receptors and their associated SMAD proteins in Caenorhabditis elegans. Development 126, 251-260. Abstract
Kuno, K., Okada, Y., Kawashima, H., Nakamura, H., Miyasaka, M., Ohno, H., and Matsushima, K. (2000). ADAMTS-1 cleaves a cartilage proteoglycan, aggrecan. FEBS Lett. 478, 241-245. Abstract Article
Kurozumi, K., Nishita, M., Yamaguchi, K., Fujita, T., Ueno, N., and Shibuya, H. (1998). BRAM1, a BMP receptor-associated molecule involved in BMP signalling. Genes Cells 3, 257-264. Abstract Article
Kurz, C.L., and Tan, M.W. (2004). Regulation of aging and innate immunity in C. elegans. Aging Cell 3, 185-193. Abstract Article
L'Etoile, N.D., Coburn, C.M., Eastham, J., Kistler, A., Gallegos, G., and Bargmann, C.I. (2002). The cyclic GMP-dependent protein kinase EGL-4 regulates olfactory adaptation in C. elegans. Neuron 36, 1079-1089. Abstract Article
Lackner, M.R., Nurrish, S.J., and Kaplan, J.M. (1999). Facilitation of synaptic transmission by EGL-30 Gqα and EGL-8 PLCβ: DAG binding to UNC-13 is required to stimulate acetylcholine release. Neuron 24, 335-346. Abstract Article
Lamoureux, F., Baud'huin, M., Duplomb, L., Heymann, D., and Redini, F. (2007). Proteoglycans: key partners in bone cell biology. Bioessays 29, 758-771. Abstract Article
Lanjuin, A., and Sengupta, P. (2002). Regulation of chemosensory receptor expression and sensory signaling by the KIN-29 Ser/Thr kinase. Neuron 33, 369-381. Abstract Article
Larsen, P.L., Albert, P.S., and Riddle, D.L. (1995). Genes that regulate both development and longevity in Caenorhabditis elegans. Genetics 139, 1567-1583. Abstract
Le Goff, C., Morice-Picard, F., Dagoneau, N., Wang, L.W., Perrot, C., Crow, Y.J., Bauer, F., Flori, E., Prost-Squarcioni, C., Krakow, D., et al. (2008). ADAMTSL2 mutations in geleophysic dysplasia demonstrate a role for ADAMTS-like proteins in TGF-β bioavailability regulation. Nat. Genet. 40, 1119-1123. Abstract Article
Lesch, B.J., and Bargmann, C.I. (2010). The homeodomain protein hmbx-1 maintains asymmetric gene expression in adult C. elegans olfactory neurons. Genes Dev. 24, 1802-1815. Abstract Article
Levy, D.E., and Darnell, J.E., Jr. (2002). Stats: transcriptional control and biological impact. Nat. Rev. Mol. Cell Biol. 3, 651-662. Abstract Article
Li, C. and Kim, K. Neuropeptides (September 25, 2008), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.142.1, http://www.wormbook.org. Article
Liang, J., Lints, R., Foehr, M.L., Tokarz, R., Yu, L., Emmons, S.W., Liu, J., and Savage-Dunn, C. (2003). The Caenorhabditis elegans schnurri homolog sma-9 mediates stage- and cell type-specific responses to DBL-1 BMP-related signaling. Development 130, 6453-6464. Abstract Article
Liang, J., Yu, L., Yin, J., and Savage-Dunn, C. (2007). Transcriptional repressor and activator activities of SMA-9 contribute differentially to BMP-related signaling outputs. Dev. Biol. 305, 714-725. Abstract Article
Lints, R., and Emmons, S.W. (1999). Patterning of dopaminergic neurotransmitter identity among Caenorhabditis elegans ray sensory neurons by a TGFβ family signaling pathway and a Hox gene. Development 126, 5819-5831. Abstract
Lints, R., and Emmons, S.W. (2002). Regulation of sex-specific differentiation and mating behavior in C. elegans by a new member of the DM domain transcription factor family. Genes Dev. 16, 2390-2402. Abstract Article
Lints, R., Jia, L., Kim, K., Li, C., and Emmons, S.W. (2004). Axial patterning of C. elegans male sensilla identities by selector genes. Dev. Biol. 269, 137-151. Abstract Article
Liu, T., Zimmerman, K.K., and Patterson, G.I. (2004). Regulation of signaling genes by TGFβ during entry into dauer diapause in C. elegans. BMC Dev. Biol. 4, 11. Abstract Article
Loria, P.M., Hodgkin, J., and Hobert, O. (2004). A conserved postsynaptic transmembrane protein affecting neuromuscular signaling in Caenorhabditis elegans. J. Neurosci. 24, 2191-2201. Abstract Article
Lozano, E., Saez, A.G., Flemming, A.J., Cunha, A., and Leroi, A.M. (2006). Regulation of growth by ploidy in Caenorhabditis elegans. Curr. Biol. 16, 493-498. Abstract Article
Luo, K., Stroschein, S.L., Wang, W., Chen, D., Martens, E., Zhou, S., and Zhou, Q. (1999). The Ski oncoprotein interacts with the Smad proteins to repress TGFβ signaling. Genes Dev. 13, 2196-2206. Abstract Article
Luo, S., Kleemann, G.A., Ashraf, J.M., Shaw, W.M., and Murphy, C.T. (2010). TGF-β and insulin signaling regulate reproductive aging via oocyte and germline quality maintenance. Cell 143, 299-312. Abstract Article
Luo, S., Shaw, W.M., Ashraf, J., and Murphy, C.T. (2009). TGF-β Sma/Mab signaling mutations uncouple reproductive aging from somatic aging. PLoS Genet. 5, e1000789. Abstract Article
MacNeil, L.T., Hardy, W.R., Pawson, T., Wrana, J.L., and Culotti, J.G. (2009). UNC-129 regulates the balance between UNC-40 dependent and independent UNC-5 signaling pathways. Nat. Neurosci. 12, 150-155. Abstract Article
Maduzia, L.L., Gumienny, T.L., Zimmerman, C.M., Wang, H., Shetgiri, P., Krishna, S., Roberts, A.F., and Padgett, R.W. (2002). lon-1 regulates Caenorhabditis elegans body size downstream of the dbl-1 TGF β signaling pathway. Dev. Biol. 246, 418-428. Abstract Article
Maduzia, L.L., Roberts, A.F., Wang, H., Lin, X., Chin, L.J., Zimmerman, C.M., Cohen, S., Feng, X.H., and Padgett, R.W. (2005). C. elegans serine-threonine kinase KIN-29 modulates TGFβ signaling and regulates body size formation. BMC Dev. Biol. 5, 8. Abstract Article
Mallo, G.V., Kurz, C.L., Couillault, C., Pujol, N., Granjeaud, S., Kohara, Y., and Ewbank, J.J. (2002). Inducible antibacterial defense system in C. elegans. Curr. Biol. 12, 1209-1214. Abstract Article
Malone, E.A., and Thomas, J.H. (1994). A screen for nonconditional dauer-constitutive mutations in Caenorhabditis elegans. Genetics 136, 879-886. Abstract
Maniere, X., Lebois, F., Matic, I., Ladoux, B., Di Meglio, J.M., and Hersen, P. (2011). Running worms: C. elegans self-sorting by electrotaxis. PLoS One 6, e16637. Abstract Article
Manning, G. Genomic overview of protein kinases (December 13, 2005), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.60.1, http://www.wormbook.org. Article
Massagué, J., Seoane, J., and Wotton, D. (2005). Smad transcription factors. Genes Dev. 19, 2783-2810. Abstract Article
McCarroll, S.A., Li, H., and Bargmann, C.I. (2005). Identification of transcriptional regulatory elements in chemosensory receptor genes by probabilistic segmentation. Curr. Biol. 15, 347-352. Abstract Article
McCulloch, D., and Gems, D. (2003). Body size, insulin/IGF signaling and aging in the nematode Caenorhabditis elegans. Exp. Gerontol. 38, 129-136. Abstract Article
McDowell, N., and Gurdon, J.B. (1999). Activin as a morphogen in Xenopus mesoderm induction. Semin. Cell Dev. Biol. 10, 311-317. Abstract Article
McEwen, J.M., Madison, J.M., Dybbs, M., and Kaplan, J.M. (2006). Antagonistic regulation of synaptic vesicle priming by Tomosyn and UNC-13. Neuron 51, 303-315. Abstract Article
McGovern, M., Yu, L., Kosinski, M., Greenstein, D., and Savage-Dunn, C. (2007). A role for sperm in regulation of egg-laying in the nematode C. elegans. BMC Dev. Biol. 7, 41. Abstract Article
McSorley, H.J., Grainger, J.R., Harcus, Y., Murray, J., Nisbet, A.J., Knox, D.P., and Maizels, R.M. (2009). daf-7-related TGF-β homologues from Trichostrongyloid nematodes show contrasting life-cycle expression patterns. Parasitology 137, 159-171. Abstract Article
Merz, D.C., Alves, G., Kawano, T., Zheng, H., and Culotti, J.G. (2003). UNC-52/perlecan affects gonadal leader cell migrations in C. elegans hermaphrodites through alterations in growth factor signaling. Dev. Biol. 256, 173-186. Abstract Article
Miller, K.G., Emerson, M.D., and Rand, J.B. (1999). Goα and diacylglycerol kinase negatively regulate the Gqα pathway in C. elegans. Neuron 24, 323-333. Abstract Article
Miller, L.M., Gallegos, M.E., Morisseau, B.A., and Kim, S.K. (1993). lin-31, a Caenorhabditis elegans HNF-3/fork head transcription factor homolog, specifies three alternative cell fates in vulval development. Genes Dev. 7, 933-947. Abstract Article
Mochii, M., Yoshida, S., Morita, K., Kohara, Y., and Ueno, N. (1999). Identification of transforming growth factor-β- regulated genes in Caenorhabditis elegans by differential hybridization of arrayed cDNAs. Proc. Natl. Acad. Sci. U. S. A. 96, 15020-15025. Abstract Article
Mörck, C., Axäng, C., and Pilon, M. (2003). A genetic analysis of axon guidance in the C. elegans pharynx. Dev. Biol. 260, 158-175. Abstract Article
Mörck, C., Vivekanand, V., Jafari, G., and Pilon, M. (2010). C. elegans ten-1 is synthetic lethal with mutations in cytoskeleton regulators, and enhances many axon guidance defective mutants. BMC Dev. Biol. 10, 55. Abstract Article
Morita, K., Chow, K.L., and Ueno, N. (1999). Regulation of body length and male tail ray pattern formation of Caenorhabditis elegans by a member of TGF-β family. Development 126, 1337-1347. Abstract
Morita, K., Flemming, A.J., Sugihara, Y., Mochii, M., Suzuki, Y., Yoshida, S., Wood, W.B., Kohara, Y., Leroi, A.M., and Ueno, N. (2002). A Caenorhabditis elegans TGF-β, DBL-1, controls the expression of LON-1, a PR-related protein, that regulates polyploidization and body length. EMBO J. 21, 1063-1073. Abstract Article
Morita, K., Shimizu, M., Shibuya, H., and Ueno, N. (2001). A DAF-1-binding protein BRA-1 is a negative regulator of DAF-7 TGF-β signaling. Proc. Natl. Acad. Sci. U. S. A. 98, 6284-6288. Abstract Article
Nagamatsu, Y., and Ohshima, Y. (2004). Mechanisms for the control of body size by a G-kinase and a downstream TGFβ signal pathway in Caenorhabditis elegans. Genes Cells 9, 39-47. Abstract Article
Nakano, Y., Nagamatsu, Y., and Ohshima, Y. (2004). cGMP and a germ-line signal control body size in C. elegans through cGMP-dependent protein kinase EGL-4. Genes Cells 9, 773-779. Abstract Article
Nam, S., Jin, Y.H., Li, Q.L., Lee, K.Y., Jeong, G.B., Ito, Y., Lee, J., and Bae, S.C. (2002). Expression pattern, regulation, and biological role of runt domain transcription factor, run, in Caenorhabditis elegans. Mol. Cell. Biol. 22, 547-554. Abstract Article
Narasimhan, S.D., Yen, K., Bansal, A., Kwon, E.S., Padmanabhan, S., and Tissenbaum, H.A. (2011). PDP-1 links the TGF-β and IIS pathways to regulate longevity, development, and metabolism. PLoS Genet. 7, e1001377. Abstract Article
Nash, B., Colavita, A., Zheng, H., Roy, P.J., and Culotti, J.G. (2000). The forkhead transcription factor UNC-130 is required for the graded spatial expression of the UNC-129 TGF-β guidance factor in C. elegans. Genes Dev. 14, 2486-2500. Abstract Article
Nelson, M.D., Zhou, E., Kiontke, K., Fradin, H., Maldonado, G., Martin, D., Shah, K., and Fitch, D.H. (2011). A bow-tie genetic architecture for morphogenesis suggested by a genome-wide RNAi screen in Caenorhabditis elegans. PLoS Genet. 7, e1002010. Abstract Article
Newfeld, S.J., Wisotzkey, R.G., and Kumar, S. (1999). Molecular evolution of a developmental pathway: phylogenetic analyses of transforming growth factor-β family ligands, receptors and Smad signal transducers. Genetics 152, 783-795. Abstract
Nicolás, F.J., De Bosscher, K., Schmierer, B., and Hill, C.S. (2004). Analysis of Smad nucleocytoplasmic shuttling in living cells. J. Cell Sci. 117, 4113-4125. Abstract Article
Nolan, K.M., Sarafi-Reinach, T.R., Horne, J.G., Saffer, A.M., and Sengupta, P. (2002). The DAF-7 TGF-β signaling pathway regulates chemosensory receptor gene expression in C. elegans. Genes Dev. 16, 3061-3073. Abstract Article
Nyström, J., Shen, Z.Z., Aili, M., Flemming, A.J., Leroi, A., and Tuck, S. (2002). Increased or decreased levels of Caenorhabditis elegans lon-3, a gene encoding a collagen, cause reciprocal changes in body length. Genetics 161, 83-97. Abstract
Oldham, S., Bohni, R., Stocker, H., Brogiolo, W., and Hafen, E. (2000). Genetic control of size in Drosophila. Philos. Trans. R. Soc., B 355, 945-952. Abstract Article
Ouellet, J., Li, S., and Roy, R. (2008). Notch signalling is required for both dauer maintenance and recovery in C. elegans. Development 135, 2583-2592. Abstract Article
Padgett, R.W., Das, P., and Krishna, S. (1998). TGF-β signaling, Smads, and tumor suppressors. Bioessays 20, 382-390. Abstract Article
Padua, D., and Massagué, J. (2009). Roles of TGFβ in metastasis. Cell Res. 19, 89-102. Abstract Article
Park, D., Estevez, A., and Riddle, D.L. (2010). Antagonistic Smad transcription factors control the dauer/non-dauer switch in C. elegans. Development 137, 477-485. Abstract Article
Park, D., Jones, K.L., Lee, H., Snutch, T.P., Taubert, S., and Riddle, D.L. (2012a). Repression of a potassium channel by nuclear hormone receptor and TGF-β signaling modulates insulin signaling in Caenorhabditis elegans. PLoS Genet. 8, e1002519. Abstract Article
Park, D., O'Doherty, I., Somvanshi, R.K., Bethke, A., Schroeder, F.C., Kumar, U., and Riddle, D.L. (2012b). Interaction of structure-specific and promiscuous G-protein-coupled receptors mediates small-molecule signaling in Caenorhabditis elegans. Proc. Natl. Acad. Sci. U. S. A. 109, 9917-9922. Article
Patel, M.N., Knight, C.G., Karageorgi, C., and Leroi, A.M. (2002). Evolution of germ-line signals that regulate growth and aging in nematodes. Proc. Natl. Acad. Sci. U. S. A. 99, 769-774. Abstract Article
Patterson, G.I., Koweek, A., Wong, A., Liu, Y., and Ruvkun, G. (1997). The DAF-3 Smad protein antagonizes TGF-β-related receptor signaling in the Caenorhabditis elegans dauer pathway. Genes Dev. 11, 2679-2690. Abstract Article
Patterson, G.I., and Padgett, R.W. (2000). TGF β-related pathways. Roles in Caenorhabditis elegans development. Trends Genet. 16, 27-33. Abstract Article
Pires-daSilva, A., and Sommer, R.J. (2003). The evolution of signalling pathways in animal development. Nat. Rev. Genet. 4, 39-49. Abstract Article
Pocock, R., Mione, M., Hussain, S., Maxwell, S., Pontecorvi, M., Aslam, S., Gerrelli, D., Sowden, J.C., and Woollard, A. (2008). Neuronal function of Tbx20 conserved from nematodes to vertebrates. Dev. Biol. 317, 671-685. Abstract Article
Podos, S.D., and Ferguson, E.L. (1999). Morphogen gradients: new insights from DPP. Trends Genet. 15, 396-402. Abstract Article
Portal-Celhay, C., Bradley, E.R., and Blaser, M.J. (2012). Control of intestinal bacterial proliferation in regulation of lifespan in Caenorhabditis elegans. BMC Microbiol. 12, 49. Abstract Article
Pyrowolakis, G., Hartmann, B., Muller, B., Basler, K., and Affolter, M. (2004). A simple molecular complex mediates widespread BMP-induced repression during Drosophila development. Dev. Cell 7, 229-240. Abstract Article
Qin, H., and Powell-Coffman, J.A. (2004). The Caenorhabditis elegans aryl hydrocarbon receptor, AHR-1, regulates neuronal development. Dev. Biol. 270, 64-75. Abstract Article
Raizen, D.M., Cullison, K.M., Pack, A.I., and Sundaram, M.V. (2006). A novel gain-of-function mutant of the cyclic GMP-dependent protein kinase egl-4 affects multiple physiological processes in Caenorhabditis elegans. Genetics 173, 177-187. Abstract Article
Rand, J.B. Acetylcholine (January 30, 2007), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.131.1, http://www.wormbook.org. Article
Rauthan, M., Morck, C., and Pilon, M. (2007). The C. elegans M3 neuron guides the growth cone of its sister cell M2 via the Kruppel-like zinc finger protein MNM-2. Dev. Biol. 311, 185-199. Abstract Article
Reece-Hoyes, J.S., Shingles, J., Dupuy, D., Grove, C.A., Walhout, A.J., Vidal, M., and Hope, I.A. (2007). Insight into transcription factor gene duplication from Caenorhabditis elegans Promoterome-driven expression patterns. BMC Genomics 8, 27. Abstract Article
Reiner, D.J., Ailion, M., Thomas, J.H., and Meyer, B.J. (2008). C. elegans anaplastic lymphoma kinase ortholog SCD-2 controls dauer formation by modulating TGF-β signaling. Curr. Biol. 18, 1101-1109. Abstract Article
Ren, P., Lim, C.S., Johnsen, R., Albert, P.S., Pilgrim, D., and Riddle, D.L. (1996). Control of C. elegans larval development by neuronal expression of a TGF-β homolog. Science 274, 1389-1391. Abstract Article
Riddle, D.L., Swanson, M.M., and Albert, P.S. (1981). Interacting genes in nematode dauer larva formation. Nature 290, 668-671. Abstract Article
Roberts, A.B., and Wakefield, L.M. (2003). The two faces of transforming growth factor β in carcinogenesis. Proc. Natl. Acad. Sci. U. S. A. 100, 8621-8623. Abstract Article
Roberts, A.F., Gumienny, T.L., Gleason, R.J., Wang, H., and Padgett, R.W. (2010). Regulation of genes affecting body size and innate immunity by the DBL-1/BMP-like pathway in Caenorhabditis elegans. BMC Dev. Biol. 10, 61. Abstract Article
Roy, P.J., Zheng, H., Warren, C.E., and Culotti, J.G. (2000). mab-20 encodes Semaphorin-2a and is required to prevent ectopic cell contacts during epidermal morphogenesis in Caenorhabditis elegans. Development 127, 755-767. Abstract
Savage, C., Das, P., Finelli, A.L., Townsend, S.R., Sun, C.Y., Baird, S.E., and Padgett, R.W. (1996). Caenorhabditis elegans genes sma-2, sma-3, and sma-4 define a conserved family of transforming growth factor β pathway components. Proc. Natl. Acad. Sci. U. S. A. 93, 790-794. Abstract Article
Savage-Dunn, C. TGF-β signaling (September 9, 2005), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.22.1, http://www.wormbook.org. Article
Savage-Dunn, C., Maduzia, L.L., Zimmerman, C.M., Roberts, A.F., Cohen, S., Tokarz, R., and Padgett, R.W. (2003). Genetic screen for small body size mutants in C. elegans reveals many TGFβ pathway components. Genesis 35, 239-247. Abstract Article
Savage-Dunn, C., Tokarz, R., Wang, H., Cohen, S., Giannikas, C., and Padgett, R.W. (2000). SMA-3 smad has specific and critical functions in DBL-1/SMA-6 TGFβ-related signaling. Dev. Biol. 223, 70-76. Abstract Article
Savage-Dunn, C., Yu, L., Gill, K., Awan, M., and Fernando, T. (2011). Non-stringent tissue-source requirements for BMP ligand expression in regulation of body size in Caenorhabditis elegans. Genet. Res. (Camb) 93, 427-432. Abstract Article
Schackwitz, W.S., Inoue, T., and Thomas, J.H. (1996). Chemosensory neurons function in parallel to mediate a pheromone response in C. elegans. Neuron 17, 719-728. Abstract Article
Schafer, W. R. Egg-laying (December 14, 2005), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.38.1, http://www.wormbook.org. Article
Schmelzle, T., and Hall, M.N. (2000). TOR, a central controller of cell growth. Cell 103, 253-262. Abstract Article
Schmierer, B., and Hill, C.S. (2007). TGFβ-SMAD signal transduction: molecular specificity and functional flexibility. Nat. Rev. Mol. Cell. Biol. 8, 970-982. Abstract Article
Schulenburg, H., Kurz, C.L., and Ewbank, J.J. (2004). Evolution of the innate immune system: the worm perspective. Immunol. Rev. 198, 36-58. Abstract Article
Sekelsky, J.J., Newfeld, S.J., Raftery, L.A., Chartoff, E.H., and Gelbart, W.M. (1995). Genetic characterization and cloning of mothers against dpp, a gene required for decapentaplegic function in Drosophila melanogaster. Genetics 139, 1347-1358. Abstract
Selvamurugan, N., Kwok, S., and Partridge, N.C. (2004). Smad3 interacts with JunB and Cbfa1/Runx2 for transforming growth factor-β1-stimulated collagenase-3 expression in human breast cancer cells. J. Biol. Chem. 279, 27764-27773. Abstract Article
Serra, R., and Chang, C. (2003). TGF-β signaling in human skeletal and patterning disorders. Birth Defects Research Part C: Embryo Today: Reviews 69, 333-351. Abstract Article
Shaw, W.M., Luo, S., Landis, J., Ashraf, J., and Murphy, C.T. (2007). The C. elegans TGF-β dauer pathway regulates longevity via insulin signaling. Curr. Biol. 17, 1635-1645. Abstract Article
Shim, J., and Lee, J. (2008). Regulation of rnt-1 expression mediated by the opposing effects of BRO-1 and DBL-1 in the nematode Caenorhabditis elegans. Biochem. Biophys. Res. Commun. 367, 130-136. Abstract Article
Siehr, M.S., Koo, P.K., Sherlekar, A.L., Bian, X., Bunkers, M.R., Miller, R.M., Portman, D.S., and Lints, R. (2011). Multiple doublesex-related genes specify critical cell fates in a C. elegans male neural circuit. PLoS One 6, e26811. Abstract Article
Singaravelu, G., Song, H.O., Ji, Y.J., Jee, C., Park, B.J., and Ahnn, J. (2007). Calcineurin interacts with KIN-29, a Ser/Thr kinase, in Caenorhabditis elegans. Biochem. Biophys. Res. Commun. 352, 29-35. Abstract Article
Snow, M.I., and Larsen, P.L. (2000). Structure and expression of daf-12: a nuclear hormone receptor with three isoforms that are involved in development and aging in Caenorhabditis elegans. Biochim. Biophys. Acta 1494, 104-116. Abstract Article
So, S., Tokumaru, T., Miyahara, K., and Ohshima, Y. (2011). Control of lifespan by food bacteria, nutrient limitation and pathogenicity of food in C. elegans. Mech. Ageing Dev. 132, 210-212. Abstract Article
Somerville, R.P., Longpre, J.M., Jungers, K.A., Engle, J.M., Ross, M., Evanko, S., Wight, T.N., Leduc, R., and Apte, S.S. (2003). Characterization of ADAMTS-9 and ADAMTS-20 as a distinct ADAMTS subfamily related to Caenorhabditis elegans GON-1. J. Biol. Chem. 278, 9503-9513. Abstract Article
Sternberg, P.W., Vulval development (June, 25 2005), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.6.1, http://www.wormbook.org. Article
Stoyanov, C.N., Fleischmann, M., Suzuki, Y., Tapparel, N., Gautron, F., Streit, A., Wood, W.B., and Muller, F. (2003). Expression of the C. elegans labial orthologue ceh-13 during male tail morphogenesis. Dev. Biol. 259, 137-149. Abstract Article
Sugawara, K., Morita, K., Ueno, N., and Shibuya, H. (2001). BIP, a BRAM-interacting protein involved in TGF-β signalling, regulates body length in Caenorhabditis elegans. Genes Cells 6, 599-606. Abstract Article
Sulston, J.E., Albertson, D.G., and Thomson, J.N. (1980). The Caenorhabditis elegans male: postembryonic development of nongonadal structures. Dev. Biol. 78, 542-576. Abstract Article
Sulston, J.E., and Horvitz, H.R. (1977). Post-embryonic cell lineages of the nematode, Caenorhabditis elegans. Dev. Biol. 56, 110-156. Abstract Article
Sun, Y., Liu, X., Eaton, E.N., Lane, W.S., Lodish, H.F., and Weinberg, R.A. (1999). Interaction of the Ski oncoprotein with Smad3 regulates TGF-β signaling. Mol. Cell 4, 499-509. Abstract Article
Suzuki, H., Yagi, K., Kondo, M., Kato, M., Miyazono, K., and Miyazawa, K. (2004). c-Ski inhibits the TGF-β signaling pathway through stabilization of inactive Smad complexes on Smad-binding elements. Oncogene 23, 5068-5076. Abstract Article
Suzuki, Y., Morris, G.A., Han, M., and Wood, W.B. (2002). A cuticle collagen encoded by the lon-3 gene may be a target of TGF-β signaling in determining Caenorhabditis elegans body shape. Genetics 162, 1631-1639. Abstract
Suzuki, Y., Yandell, M.D., Roy, P.J., Krishna, S., Savage-Dunn, C., Ross, R.M., Padgett, R.W., and Wood, W.B. (1999). A BMP homolog acts as a dose-dependent regulator of body size and male tail patterning in Caenorhabditis elegans. Development 126, 241-250. Abstract
Swanson, M.M., and Riddle, D.L. (1981). Critical periods in the development of the Caenorhabditis elegans dauer larva. Dev. Biol. 84, 27-40. Abstract Article
ten Dijke, P., and Hill, C.S. (2004). New insights into TGF-β-Smad signalling. Trends Biochem. Sci. 29, 265-273. Abstract Article
Tenor, J.L., and Aballay, A. (2008). A conserved Toll-like receptor is required for Caenorhabditis elegans innate immunity. EMBO Rep. 9, 103-109. Abstract Article
Tewari, M., Hu, P.J., Ahn, J.S., Ayivi-Guedehoussou, N., Vidalain, P.O., Li, S., Milstein, S., Armstrong, C.M., Boxem, M., Butler, M.D., et al. (2004). Systematic interactome mapping and genetic perturbation analysis of a C. elegans TGF-β signaling network. Mol. Cell 13, 469-482. Abstract Article
Thomas, J.H., Birnby, D.A., and Vowels, J.J. (1993). Evidence for parallel processing of sensory information controlling dauer formation in Caenorhabditis elegans. Genetics 134, 1105-1117. Abstract
Tian, C., Sen, D., Shi, H., Foehr, M.L., Plavskin, Y., Vatamaniuk, O.K., and Liu, J. (2010). The RGM protein DRAG-1 positively regulates a BMP-like signaling pathway in Caenorhabditis elegans. Development 137, 2375-2384. Abstract Article
Tiedemann, K., Sasaki, T., Gustafsson, E., Göhring, W., Batge, B., Notbohm, H., Timpl, R., Wedel, T., Schlötzer-Schrehardt, U., and Reinhardt, D.P. (2005). Microfibrils at basement membrane zones interact with perlecan via fibrillin-1. J. Biol. Chem. 280, 11404-11412. Abstract Article
Tissenbaum H.A., Hawdon J., Perregaux M., Hotez P., Guarente L., and Ruvkun G. (2000). A common muscarinic pathway for diapause recovery in the distantly related nematode species Caenorhabditis elegans and Ancylostoma caninum. Proc. Natl. Acad. Sci. U. S. A. 97, 460-465. Abstract Article
Tortorella, M., Pratta, M., Liu, R.Q., Abbaszade, I., Ross, H., Burn, T., and Arner, E. (2000). The thrombospondin motif of aggrecanase-1 (ADAMTS-4) is critical for aggrecan substrate recognition and cleavage. J. Biol. Chem. 275, 25791-25797. Abstract Article
Tortorella, M.D., Arner, E.C., Hills, R., Gormley, J., Fok, K., Pegg, L., Munie, G., and Malfait, A.M. (2005). ADAMTS-4 (aggrecanase-1): N-terminal activation mechanisms. Arch. Biochem. Biophys. 444, 34-44. Abstract Article
Trent, C., Tsuing, N., and Horvitz, H.R. (1983). Egg-laying defective mutants of the nematode Caenorhabditis elegans. Genetics 104, 619-647. Abstract
van der Linden, A.M., Wiener, S., You, Y.J., Kim, K., Avery, L., and Sengupta, P. (2008). The EGL-4 PKG acts with KIN-29 salt-inducible kinase and protein kinase A to regulate chemoreceptor gene expression and sensory behaviors in Caenorhabditis elegans. Genetics 180, 1475-1491. Abstract Article
Vashlishan, A.B., Madison, J.M., Dybbs, M., Bai, J., Sieburth, D., Ch'ng, Q., Tavazoie, M., and Kaplan, J.M. (2008). An RNAi screen identifies genes that regulate GABA synapses. Neuron 58, 346-361. Abstract Article
Velasco, J., Li, J., DiPietro, L., Stepp, M.A., Sandy, J.D., and Plaas, A. (2011). Adamts5 deletion blocks murine dermal repair through CD44-mediated aggrecan accumulation and modulation of transforming growth factor β1 (TGFβ1) signaling. J. Biol. Chem. 286, 26016-26027. Abstract Article
Viney, M.E., Thompson, F.J., and Crook, M. (2005). TGF-β and the evolution of nematode parasitism. Int. J. Parasitol. 35, 1473-1475. Abstract Article
Wang, J., Farr, G.W., Hall, D.H., Li, F., Furtak, K., Dreier, L., and Horwich, A.L. (2009). An ALS-linked mutant SOD1 produces a locomotor defect associated with aggregation and synaptic dysfunction when expressed in neurons of Caenorhabditis elegans. PLoS Genet. 5, e1000350. Abstract Article
Wang, J., Mohler, W.A., and Savage-Dunn, C. (2005). C-terminal mutants of C. elegans Smads reveal tissue-specific requirements for protein activation by TGF-β signaling. Development 132, 3505-3513. Abstract Article
Wang, J., Tokarz, R., and Savage-Dunn, C. (2002). The expression of TGFβ signal transducers in the hypodermis regulates body size in C. elegans. Development 129, 4989-4998. Abstract
Wang, W., Mariani, F.V., Harland, R.M., and Luo, K. (2000). Ski represses bone morphogenic protein signaling in Xenopus and mammalian cells. Proc. Natl. Acad. Sci. U. S. A. 97, 14394-14399. Abstract Article
Wang, W.M., Lee, S., Steiglitz, B.M., Scott, I.C., Lebares, C.C., Allen, M.L., Brenner, M.C., Takahara, K., and Greenspan, D.S. (2003). Transforming growth factor-β induces secretion of activated ADAMTS-2. A procollagen III N-proteinase. J. Biol. Chem. 278, 19549-19557. Abstract Article
Wang, Y., and Levy, D.E. (2006). C. elegans STAT cooperates with DAF-7/TGF-β signaling to repress dauer formation. Curr. Biol. 16, 89-94. Abstract Article
Watanabe, N., Ishihara, T., and Ohshima, Y. (2007). Mutants carrying two sma mutations are super small in the nematode C. elegans. Genes Cells 12, 603-609. Abstract Article
Watanabe, N., Nagamatsu, Y., Gengyo-Ando, K., Mitani, S., and Ohshima, Y. (2005). Control of body size by SMA-5, a homolog of MAP kinase BMK1/ERK5, in C. elegans. Development 132, 3175-3184. Abstract Article
Wilkinson, L., Kolle, G., Wen, D., Piper, M., Scott, J., and Little, M. (2003). CRIM1 regulates the rate of processing and delivery of bone morphogenetic proteins to the cell surface. J. Biol. Chem. 278, 34181-34188. Abstract Article
Wong, Y.F., Sheng, Q., Chung, J.W., Chan, J.K., and Chow, K.L. (2010). mab-31 and the TGF-β pathway act in the ray lineage to pattern C. elegans male sensory rays. BMC Dev. Biol. 10, 82. Abstract Article
Wrana, J.L., and Attisano, L. (1996). MAD-related proteins in TGF-β signalling. Trends Genet. 12, 493-496. Abstract Article
Wu, M.Y., and Hill, C.S. (2009). TGF-β superfamily signaling in embryonic development and homeostasis. Dev. Cell 16, 329-343. Abstract Article
Xia, Y., Babitt, J.L., Sidis, Y., Chung, R.T., and Lin, H.Y. (2008). Hemojuvelin regulates hepcidin expression via a selective subset of BMP ligands and receptors independently of neogenin. Blood 111, 5195-5204. Abstract Article
Xiao, Z., Watson, N., Rodriguez, C., and Lodish, H.F. (2001). Nucleocytoplasmic shuttling of Smad1 conferred by its nuclear localization and nuclear export signals. J. Biol. Chem. 276, 39404-39410. Abstract Article
Xu, L., Kang, Y., Col, S., and Massagué, J. (2002). Smad2 nucleocytoplasmic shuttling by nucleoporins CAN/Nup214 and Nup153 feeds TGFβ signaling complexes in the cytoplasm and nucleus. Mol. Cell 10, 271-282. Abstract Article
Xu, W., Angelis, K., Danielpour, D., Haddad, M.M., Bischof, O., Campisi, J., Stavnezer, E., and Medrano, E.E. (2000). Ski acts as a co-repressor with Smad2 and Smad3 to regulate the response to type β transforming growth factor. Proc. Natl. Acad. Sci. U. S. A. 97, 5924-5929. Abstract Article
Yin, J., Yu, L., and Savage-Dunn, C. (2010). Alternative trans-splicing of Caenorhabditis elegans sma-9/schnurri generates a short transcript that provides tissue-specific function in BMP signaling. BMC Mol. Biol. 11, 46. Abstract Article
Yoshida, S., Morita, K., Mochii, M., and Ueno, N. (2001). Hypodermal expression of Caenorhabditis elegans TGF-β type I receptor SMA-6 is essential for the growth and maintenance of body length. Dev. Biol. 240, 32-45. Abstract Article
Yoshimura, A., Wakabayashi, Y., and Mori, T. (2010). Cellular and molecular basis for the regulation of inflammation by TGF-β. J. Biochem. 147, 781-792. Abstract Article
You, Y.J., Kim, J., Raizen, D.M., and Avery, L. (2008). Insulin, cGMP, and TGF-β signals regulate food intake and quiescence in C. elegans: a model for satiety. Cell Metab. 7, 249-257. Abstract Article
Zhang, X., and Zhang, Y. (2009). Neural-immune communication in Caenorhabditis elegans. Cell Host Microbe 5, 425-429. Abstract Article
Zhang, Y.W., Yasui, N., Ito, K., Huang, G., Fujii, M., Hanai, J., Nogami, H., Ochi, T., Miyazono, K., and Ito, Y. (2000). A RUNX2/PEBP2α A/CBFA1 mutation displaying impaired transactivation and Smad interaction in cleidocranial dysplasia. Proc. Natl. Acad. Sci. U. S. A. 97, 10549-10554. Abstract Article
*Edited by Iva Greenwald. Last revised July 23, 2012, Published July 10, 2013. This chapter should be cited as: Gumienny T.L., Savage-Dunn C. TGF-β signaling in C. elegans (July 10, 2013), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.22.2, http://www.wormbook.org.
Copyright: © 2013 Tina L. Gumienny and Cathy Savage-Dunn. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
§To whom correspondence should be addressed. E-mail: Cathy.SavageDunn@qc.cuny.edu
 All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.
All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.