Embryo series courtesy of Einhard Schierenberg
Embryo series courtesy of Einhard SchierenbergTable of Contents
Abstract
During the first stage of larval development, the Q neuroblasts and their descendants migrate to well-defined positions along the anteroposterior body axis, where they differentiate into sensory neurons and interneurons. The two Q neuroblasts are initially present at similar positions on the left and right lateral side, but this symmetry is broken when the Q neuroblast on the left side (QL) polarizes towards the posterior and the Q neuroblast on the right side (QR) towards the anterior. This left-right asymmetry is maintained when the descendants of the two Q neuroblasts migrate to their final positions in the posterior and anterior. The mechanisms that establish this asymmetry and control the migration of the Q descendants along the anteroposterior axis are surprisingly complex and include interplay between Wnt signaling pathways, homeotic genes, and the basic cell migration and polarity machinery. Here, we will give an overview of what is currently known about the mechanisms that mediate and control the development and migration of the Q neuroblasts and their descendants.
During late embryogenesis and early larval development, several neuronal cells migrate along the anteroposterior body axis (Sulston and Horvitz, 1977; Hedgecock et al., 1987; Silhankova and Korswagen, 2007). Among these are the Q neuroblasts and their descendants, which differentiate into distinct sensory neurons and interneurons.
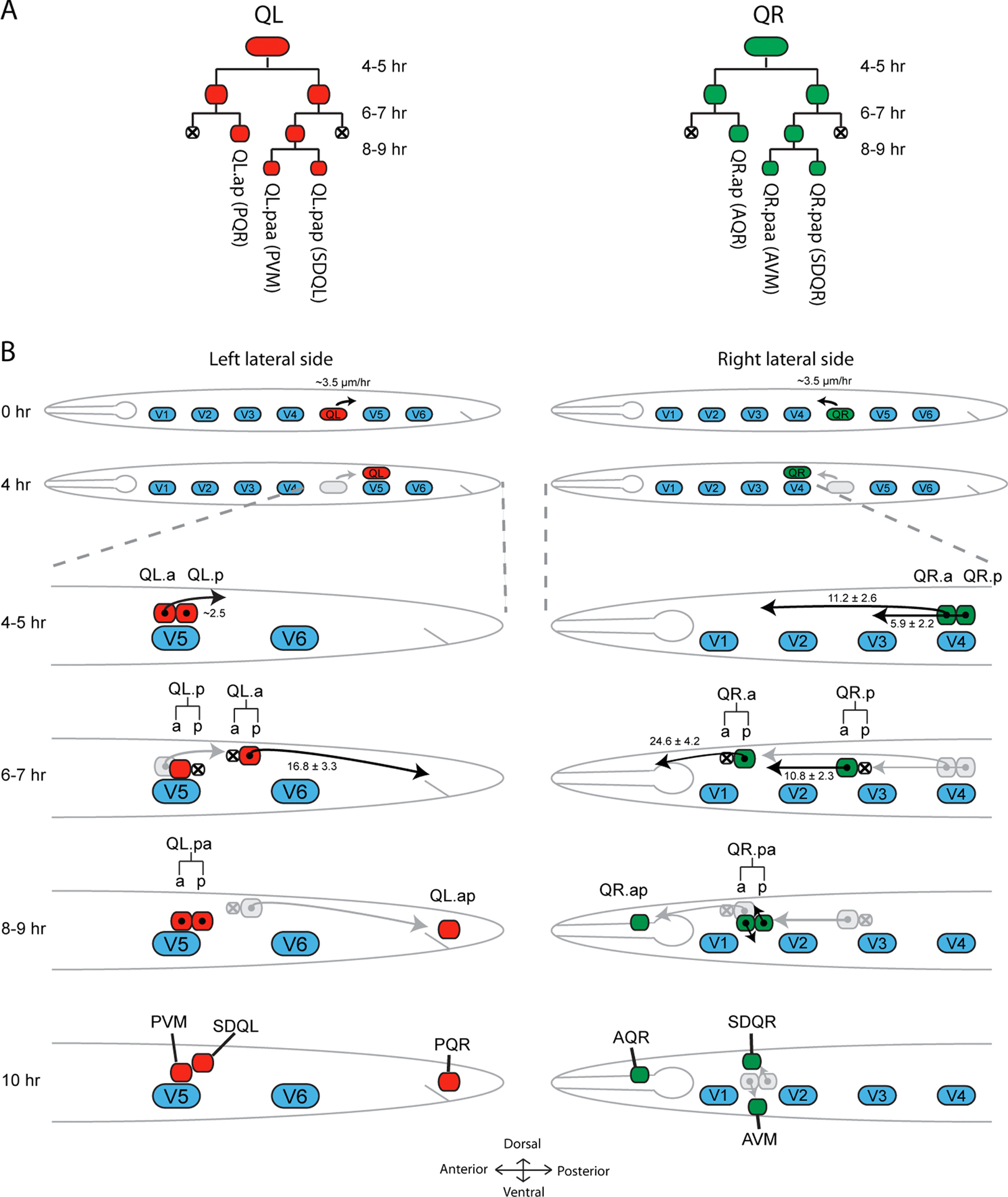
The migration of the Q neuroblasts commences when the left Q neuroblast QL and the right Q neuroblast QR—which are initially present at similar positions within the lateral row of seam cells on the left and right side of the animal—polarize and migrate in opposite directions (Figure 1, Figure 2, Movie 1, Movie 2) (Sulston and Horvitz, 1977; Honigberg and Kenyon, 2000). During this phase, QL migrates a short distance towards the posterior to a position dorsal of the seam cell V5, while QR migrates towards the anterior to a position above the seam cell V4. After this initial migration, the Q neuroblasts divide into the equally sized daughter cells Q.a and Q.p that migrate further along the anteroposterior axis. During this migration, Q.a divides once and Q.p twice to generate a total of three descendants that migrate towards well-defined positions in the posterior (QL descendants) or anterior (QR descendants) to differentiate into the sensory neurons PVM and PQR, AVM and AQR, and the interneurons SDQR and SDQL (Figure 1, Figure 4, Movie 3, Movie 4) (Sulston and Horvitz, 1977; Chalfie and Sulston, 1981). In addition, two descendants (Q.aa and Q.pp) are formed that undergo apoptosis.
|
Figure 1. Q cell lineage and schematic overview of their migration. (A) Both Q neuroblasts undergo an identical pattern of symmetric and asymmetric cell divisions generating two cells that undergo apoptosis (Q.aa and Q.pp) and three cells that after completing their migration differentiate into specific sensory neurons and interneurons. (B) At the time of hatching (0 h) both Q neuroblasts are located within the two lateral rows of seam cells (V1-V6) at a position located between V4 and V5. During L1 development the Q cell on the right lateral side (QR) will migrate anteriorly while the left Q cell (QL) migrates posteriorly. After approximately 4 h, QL is positioned above V5 while QR lies above V4. This initial short range migration is followed by the first cell division (4-5 h). The Q descendants (Q.x) migrate further along the anteroposterior axis. During their long range migration, Q.a and Q.p undergo one and two additional cell divisions, respectively. Different cells in the Q lineage migrate with different velocities. Values in B indicate migration speed in μm/h (Ou and Vale, 2009). At the end of L1, three QL.x and three QR.x occupy well-defined final positions and differentiate into neurons. Body axes are as indicated.
The two phases of the migration are controlled by distinct mechanisms. The initial phase, in which the left-right asymmetry is established and the Q neuroblasts polarize and migrate a short distance in opposite directions, requires the netrin receptor UNC-40 and the transmembrane proteins MIG-21 and DPY-19 (Honigberg and Kenyon, 2000; Middelkoop et al., 2012; Sundararajan and Lundquist, 2012). This phase is terminated when the Q neuroblasts undergo their first division. In the second phase, a complex interplay between Wnt signaling pathways, homeotic genes, and the basic cell migration and polarity machinery maintains the left-right asymmetry in migration direction and guides the Q neuroblast descendants to their final positions along the anteroposterior body axis (Harris et al., 1996; Maloof et al., 1999; Whangbo and Kenyon, 1999; Honigberg and Kenyon, 2000; Zinovyeva et al., 2008).
The migration of the Q neuroblasts provides an excellent model to study various basic questions in developmental biology and cell biology. First, it is one of the few examples of left-right asymmetry in the largely bilateral C. elegans body plan (Sulston and Horvitz, 1977; Hedgecock et al., 1987). Since the Q neuroblasts are identical with respect to their initial position, morphology, and cell lineage, a key question is how this left-right asymmetry is established. Second, the guidance of neuronal precursor cells along the anteroposterior axis and the role of Wnt proteins in this process appears to be an evolutionarily conserved mechanism in nervous system development (Silhankova and Korswagen, 2007). Finally, the Q neuroblasts undergo both symmetric and asymmetric cell divisions during their migration, providing an in vivo system to study these processes in motile cells.
The migration of the Q neuroblasts has been intensively studied over the past two decades. Important insight into the role of homeotic genes in establishing left-right asymmetry in Q cell migration was gained from pioneering studies by the group of Cynthia Kenyon (Salser and Kenyon, 1992). With the development of GFP markers that label the Q neuroblast descendants in living animals (Chalfie et al., 1994), it became possible to perform large scale forward genetic screens for mutants that disrupt Q cell migration (Ch'ng et al., 2003). These screens and the subsequent analysis of the mutated gene products have generated a wealth of information on the molecular mechanisms that are involved. Here, we will give an overview of these mechanisms. We will first discuss the Q neuroblast lineage and the mechanisms that establish the initial left-right asymmetry in Q neuroblast migration. In the second part, we will discuss how the Q neuroblast descendants are guided towards their final destinations and how the two phases of Q cell migration are interconnected.
During the first stage of larval development, QL and QR undergo a similar pattern of cell divisions (Sulston and Horvitz, 1977; Hedgecock et al., 1987). The first division generates daughter cells of equal size (Q.a and Q.p), while the second division is asymmetric, with Q.a generating a small anterior daughter (Q.aa) that goes into apoptosis, and a larger posterior daughter (Q.ap) that differentiates into the sensory neuron PQR (in case of QL.ap) or AQR (QR.ap). In contrast, Q.p produces a small posterior daughter cell (Q.pp) that goes into apoptosis, while the larger anterior daughter (Q.pa) survives. Q.pa in turn undergoes a third round of symmetric cell division, generating the daughter cells Q.pap and Q.paa that differentiate into SDQL/R and P/AVM, respectively (Figure 1) (Sulston and Horvitz, 1977). The apoptotic corpses of Q.aa and Q.pp are cleared by the surrounding hypodermis, a process that is dependent on engulfment and autophagy related genes (Li et al., 2012; Zou et al., 2012).
An important regulator of the asymmetric divisions in the Q neuroblast lineage is the MELK (Maternal Embryonic Leucine zipper Kinase) ortholog, pig-1, which belongs to the PAR-1/Kin1/SAD-1 superfamily of serine/threonine kinases that regulate cell polarity in various contexts (Drewes and Nurse, 2003; La Carbona et al., 2004; Kishi et al., 2005; Cordes et al., 2006; Ou et al., 2010). PIG-1 contains an N-terminal kinase domain and a C-terminal kinase associated domain, both of which are necessary for the function of PIG-1 in asymmetric cell division (Cordes et al., 2006; Chien et al., 2013). In pig-1 loss of function mutants, the Q.a and Q.p divisions generate equally sized daughter cells. Interestingly, Q.pp frequently fails to undergo apoptosis in pig-1 mutants and adopts the Q.pa fate, resulting in extra SDQL/R and P/AVM neurons. This ectopic differentiation cannot be solely explained by the lack of apoptosis since the Q.pp cell does not typically adopt a Q.pa fate in apoptosis defective mutants. However, blocking apoptosis in a pig-1 background, or in other mutants that display asymmetric Q.p division defects (see below), dramatically enhances the extra Q.pa phenotype. Together these results suggest that the increased Q.pp cell size favors the Q.pa fate (Cordes et al., 2006). Members of the PAR-1/Kin1/SAD-1 superfamily are phosphorylated and activated by the LKB1 kinase, which is part of a complex containing the pseudokinase STRAD and MO25 (Lizcano et al., 2004). The C. elegans orthologs of LKB1 (par-4), STRAD (strd-1), and one isoform of the MO25 ortholog, mop-25, are also involved in the control of asymmetric Q.p cell division, and mutants of these genes display similar phenotypes as observed in pig-1. Genetic analysis suggests that pig-1 functions in a linear pathway with par-4 and strd-1 (Chien et al., 2013). This conclusion is supported by the observation that a conserved threonine residue in the PIG-1 kinase domain (T169), which is phosphorylated by LKB1 in other systems (Lizcano et al., 2004), is necessary for PIG-1 function in Q.p division as well (Chien et al., 2013).
Another component that is necessary for asymmetric Q.p division is encoded by the gene cnt-2 (Singhvi et al., 2011). As in pig-1 mutants, the division of Q.p is symmetric in cnt-2 mutants, and double mutant analysis suggests that both genes function in a linear pathway. cnt-2 encodes an Arf GTPase activating protein (GAP) that belongs to the AGAP family. One of the proposed targets of cnt-2 is the ADP-ribosylation factor arf-1. The Q.p lineage defect of arf-1 mutants is however less severe than in cnt-2 mutants, indicating that CNT-2 has additional targets. Interestingly, CNT-2 and ARF-1 are required for receptor mediated endocytosis and it was found that interference with endocytosis results in mild Q.p division defects (Singhvi et al., 2011). The role of endocytosis in Q.p division and its relation to the action of pig-1 remains to be established.
Although both Q.a and Q.p divide asymmetrically, generate a daughter cell that undergoes apoptosis, and require functional pig-1, the polarity of the two lineally equivalent cells is oriented oppositely. This indicates that Q.a and Q.p use different mechanisms to achieve polarity and asymmetric division. In Q.p, the mitotic spindle is displaced posteriorly from metaphase onwards, resulting in cleavage furrow formation posterior to the center of the cell and yielding a larger anterior daughter and a smaller posterior daughter. In Q.a, spindle asymmetry initiates much later, during anaphase, and this asymmetry increases progressively during mitosis (Ou et al., 2010). The non-muscle myosin II protein NMY-2 is a key component of the cell division machinery that localizes to the contractile ring of dividing Q.a and Q.p cells. In the case of Q.a, it was found that NMY-2 also localizes to the anterior cortex during division. By performing chromophore-assisted laser inactivation (CALI), Ou and colleagues (Ou et al., 2010) elegantly showed that this polarized NMY-2 distribution is essential for the asymmetric division of Q.a. The asymmetric distribution of NMY-2 likely results in the generation of strong anterior cortical forces which in turn cause asymmetric distribution of the cytoplasm and polarized spindle positioning. NMY-2 is not asymmetrically localized in Q.p, indicating that mitotic spindle orientation and asymmetric division relies on a different mechanism (Ou et al., 2010).
The left-right asymmetry in Q cell migration commences shortly after hatching when QL and QR polarize in opposite directions. This initial polarization process is followed by a short-range migration (approximately 10-15 μm in a timeframe of 4-5 h), where QL migrates posteriorly and QR migrates anteriorly (Figure 1, Figure 2, Movie 1, Movie 2) (Sulston and Horvitz, 1977). Studies that examined the first steps of Q cell migration provided evidence that initial Q neuroblast migration (prior to the first Q cell division) and Q descendant migration are controlled by different mechanisms.
|
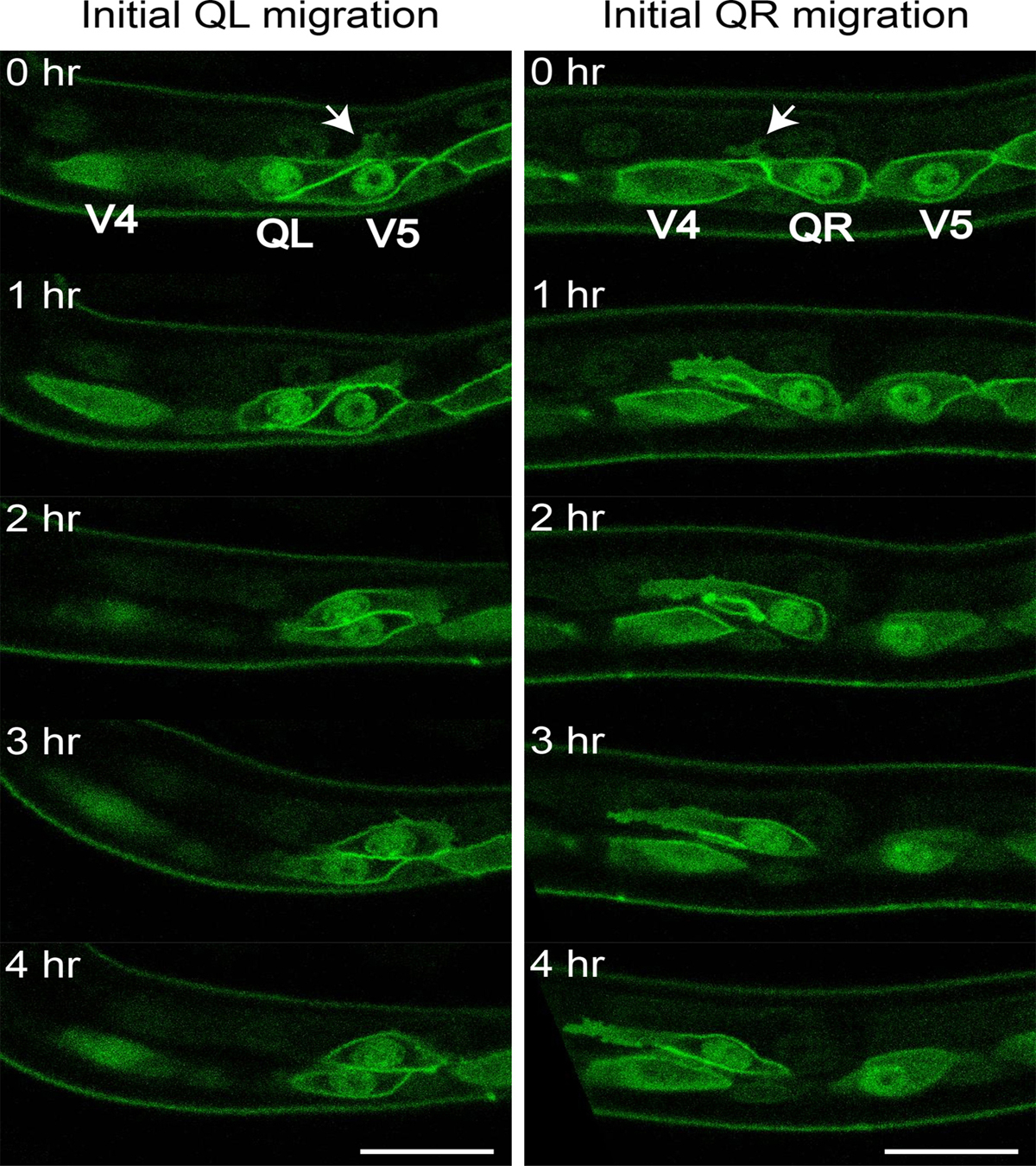
Figure 2. Initial Q cell polarization and migration. Figure shows representative still images from Movie 1 (left panels, QL) and Movie 2 (right panels, QR). Arrows mark initial Q neuroblast protrusions that will grow over time towards the posterior (QL) or the anterior (QR). Scale bars depict 10 μm.
Movie 1. Initial QL polarization and migration. Movie shows a QL neuroblast at the onset of initial migration. It has sent a posterior protrusion reaching over seam cell V5. Over time, the cell body follows the protruding front and at the end of initial migration QL is positioned above the seam cell V5. Q cells and the seam cells express GFP-PH (Pleckstrin homology domain, marking the cell periphery) and H2B::GFP (marking the nuclei) under the control of a seam and Q cell specific promoter (Wildwater et al., 2011; Middelkoop et al., 2012). Thirty min time intervals shown at 1 frame/s, dorsal is up, anterior is left. Movie is adapted from (Middelkoop et al., 2012).
Movie 2. Initial QR polarization and migration. Movie shows a QR neuroblast at the onset of initial migration. It has sent an anterior protrusion reaching over seam cell V4. Over time, the cell body follows the protruding front and at the end of initial migration QR is positioned above seam cell V4. Markers as in Movie 1. Thirty min time intervals shown at 1 frame/s, dorsal is up, anterior is left. Movie is adapted from (Middelkoop et al., 2012).
The direction of initial Q polarization and migration is randomized in unc-40 mutants. unc-40 encodes the C. elegans ortholog of DCC (Deleted in Colorectal Cancer) and is well known for its function in dorsoventral guidance of cells and axons, where it acts as a receptor for the conserved UNC-6/netrin guidance cue (Chan et al., 1996; Wadsworth, 2002). Time-lapse analysis of Q cell migration revealed that in unc-40 mutants the Q cells polarize in multiple directions, indicating that this process becomes variable in the absence of unc-40. Analysis of an UNC-40::GFP reporter gene showed that unc-40 is strongly expressed in the Q neuroblasts. Furthermore, Q cell-specific expression of unc-40 rescued the mutant phenotype, suggesting that it functions cell-autonomously (Honigberg and Kenyon, 2000; Sundararajan and Lundquist, 2012). Interestingly, unlike its well-characterized role in dorsoventral axon guidance, the function of UNC-40 in Q cell polarization neither depends on UNC-6/netrin nor on the putative co-receptor UNC-5 (Honigberg and Kenyon, 2000).
Loss of the thrombospondin repeat-containing transmembrane protein MIG-21 results in similar defects as unc-40 (Du and Chalfie, 2001; Middelkoop et al., 2012; Sundararajan and Lundquist, 2012). In mig-21 mutants, QL and QR fail to persistently polarize in one direction and repeatedly change direction during the initial migration phase. It was shown that mig-21 is transiently expressed in the Q neuroblasts during the initial migration phase and that it functions cell-autonomously (Middelkoop et al., 2012; Sundararajan and Lundquist, 2012).
Another gene involved in controlling initial Q cell migration is dpy-19 (Honigberg and Kenyon, 2000). dpy-19 encodes a predicted transmembrane protein with 13 hydrophobic regions and shows homology to four mammalian DPY-19 like proteins, mDpy19L1 to L4 (Carson et al., 2006). While unc-40 mutants have a variety of defects in other cell and axon migrations, dpy-19 only displays a Q neuroblast migration phenotype. As in unc-40 and mig-21 mutants, the direction of initial Q neuroblast polarization is random in dpy-19 mutants (Honigberg and Kenyon, 2000). The mammalian ortholog mDpy19L1 was shown to control radial migration of glutamatergic neurons, possibly pointing towards a conserved role of DPY-19 in neuronal migration (Watanabe et al., 2011). In addition, mDpy19L2 was found to localize to the inner nuclear membrane in sperm cells, where it controls acrosome maturation (Harbuz et al., 2011; Pierre et al., 2012). Whether a similar process occurs in cell migration remains to be determined. Analysis of mig-21 dpy-19 double mutants suggests that they function in a linear genetic pathway (Middelkoop et al., 2012). This conclusion is supported by the finding that DPY-19 functions as a C-mannosyltransferase that mannosylates the thrombospondin repeats of MIG-21, a modification that is necessary for the secretion of a recombinant soluble form of MIG-21 (Buettner et al., 2013). It has however not been demonstrated that this modification is also required for MIG-21 function in vivo.
In addition to dpy-19 and mig-21, loss of one of the isoforms of the LAR receptor tyrosine-like phosphatase ptp-3 also affects the initial polarization of the Q neuroblasts (Sundararajan and Lundquist, 2012). ptp-3B encodes a transmembrane protein and has an extracellular domain with two fibronectin type III repeats and an intracellular domain with two tyrosine phosphatase domains. Q neuroblast specific expression of ptp-3B rescued the ptp-3 mutant phenotype, suggesting that ptp-3 functions cell autonomously in the Q cells. Epistasis analysis suggests that unc-40 acts in parallel to dpy-19, mig-21, and ptp-3. In addition, the mig-21 single mutant phenotype was not enhanced by loss of ptp-3, indicating that ptp-3 functions in the mig-21- and dpy-19-containing branch of the initial Q cell migration pathway (Honigberg and Kenyon, 2000; Middelkoop et al., 2012; Sundararajan and Lundquist, 2012). unc-40, ptp-3, dpy-19, and mig-21 all encode transmembrane proteins that could potentially function as guidance receptors. However, to date no secreted factor has been identified that controls the initial migration of the Q neuroblasts.
In addition to the proteins described above, Chapman and colleagues found that the Nck-interacting kinase (NIK) ortholog MIG-15 also plays a role during the initial phase of Q neuroblast migration (Chapman et al., 2008). They found that mig-15 acts cell autonomously in the Q neuroblasts to control polarization and migration, a phenotype that strongly resembles that of unc-40 and dpy-19 mutants. Vertebrate NIK and the Drosophila ortholog Misshapen have been shown to act in the JNK/MAPK pathway and have been implicated in the control of cell migration (Su et al., 1998; Xue et al., 2001). Furthermore, tissue culture experiments have shown that NIK-mediated ERM phosphorylation mediates lammellipodia formation via actin reorganization (Baumgartner et al., 2006). MIG-15 contains a STE20-like serine/threonine protein kinase domain, a proline-rich domain and a Citron-NIK homology (CNH) domain. Interestingly, Citron proteins interact with Rho GTPases and are thought to function as Rho effectors during cytokinesis (Madaule et al., 2000). Given that members of the Rho/Rac family are also required for Q neuroblast migration (see below), MIG-15 could potentially function as an effector of Rho/Rac GTPases in Q cell migration. Genetic analysis indicates that the kinase domain (as well as sequences C-terminal of this domain) are necessary for MIG-15 to exert its function in Q cell migration (Shakir et al., 2006; Chapman et al., 2008). More recently, Mahmoudi and coworkers described a role for mammalian TNIK in Wnt signaling, where TNIK interacts directly with the TCF transcription factor Tcf4 to activate canonical Wnt/β-catenin signaling (Mahmoudi et al., 2009). Although mig-15 acts during initial Q polarization and migration, which is Wnt independent, a role in the subsequent Wnt dependent migration of the Q descendants cannot be excluded.
Cell motility is facilitated by the dynamic remodeling of the actin cytoskeleton. The main regulators of actin dynamics are members of the Rho/Rac-like family of small GTPases that oscillate between GTP and GDP bound states. The C. elegans genome contains three Rho/Rac-like GTPases: ced-10/Rac1, rac-2/Rac1, and mig-2/RhoG (Zipkin et al., 1997; Lundquist et al., 2001) (see WormBook chapter Small GTPases for a detailed overview of these proteins in C. elegans). Genetic analysis revealed that ced-10 and mig-2 redundantly control initial Q cell migration, with loss of both ced-10 and mig-2 severely affecting the ability of the Q cells to send out membrane protrusions and thereby impair the extent of their migration (Dyer et al., 2010).
The guanine nucleotide exchange factor (GEF) UNC-73/Trio also plays an important role in Q cell migration. Although less severe, the phenotype of unc-73 is similar to that of ced-10; mig-2 double mutants. unc-73 is expressed in the migrating Q neuroblasts and their descendants from the moment of initial neuroblast migration (Hedgecock et al., 1987; Steven et al., 1998; Honigberg and Kenyon, 2000; Dyer et al., 2010). unc-73 encodes a protein with two Dbl homology domains (DH-1 and DH-2) that are separated by a pleckstrin homology domain (PH-1), an SH3 domain, and a putative PEST sequence. Furthermore, another PH domain (PH-2), an Ig-like domain, and a fibronectin type III domain are located downstream of the DH-2 domain (Steven et al., 1998). In vitro analysis has shown that recombinant UNC-73 containing the DH-1 and PH-1 domains has GEF activity for MIG-2 and CED-10 but not for RHO-1 and CDC-42 (Wu et al., 2002; Kubiseski et al., 2003). In addition, recombinant UNC-73 containing the DH-1, PH-1, and SH3 domains can promote GDP/GTP exchange on human Rac1, but not on human Cdc42 or Rho, indicating that unc-73 encodes a well-conserved GEF that specifically activates Rac GTPases (Figure 3) (Steven et al., 1998).
|
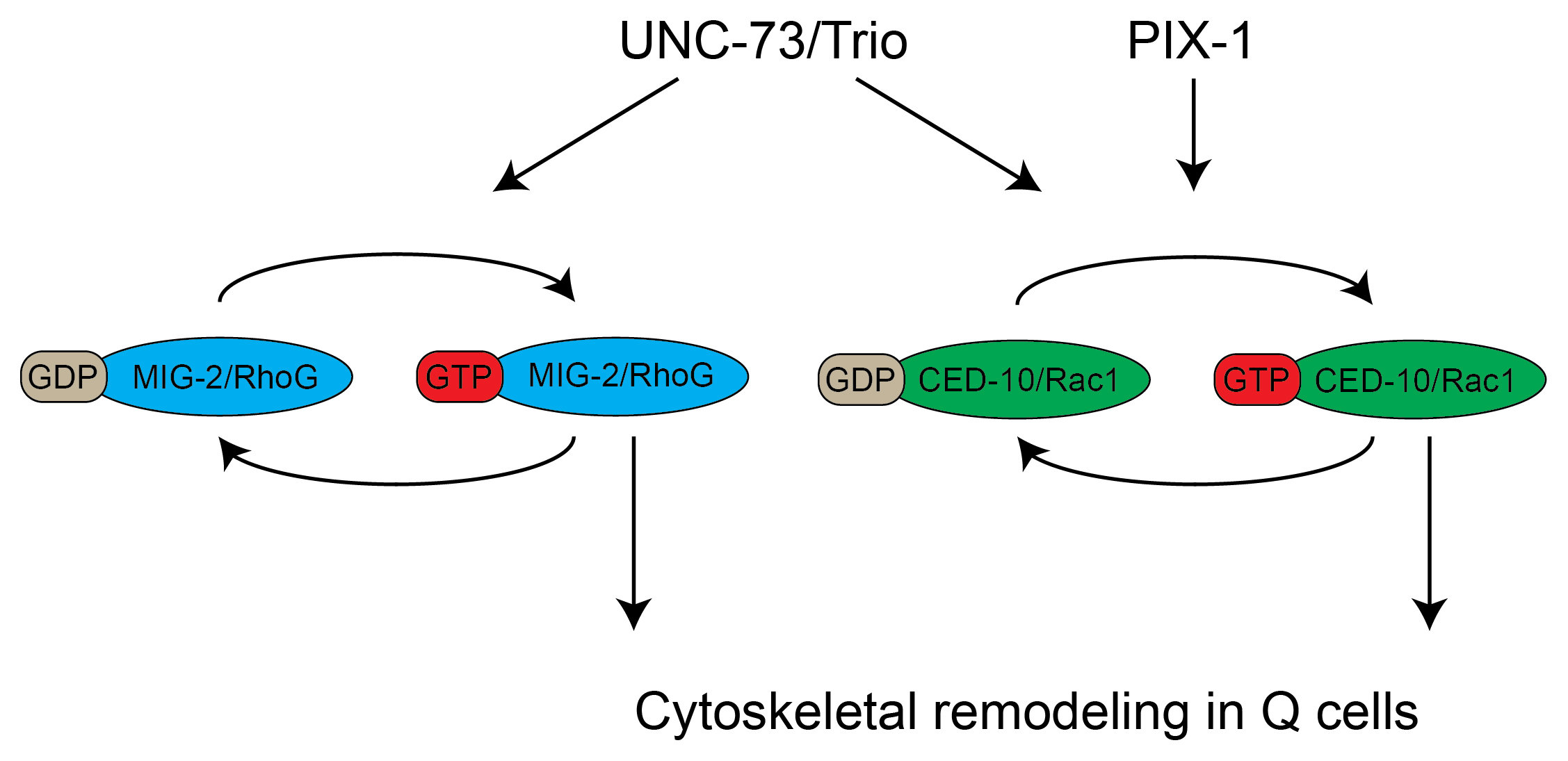
Figure 3. Two parallel pathways control cytoskeletal remodeling in migrating Q neuroblast descendants. MIG-2/RhoG and CED-10/Rac1 cycle between active, GTP bound, and inactive, GDP bound, states. Both CED-10 and MIG-2 contribute to the migration. The guanine nucleotide exchange factors UNC-73/Trio and PIX-1 were shown to act upstream of MIG-2 and CED-10. Genetic evidence suggests that PIX-1 only activates CED-10, while UNC-73 activates both MIG-2 and CED-10.
When the genetic and biochemical data are combined, it is likely that unc-73 acts as a GEF that functions upstream of mig-2 and ced-10. However, the Q cell migration phenotype of unc-73 null mutants is not as severe as that of ced-10; mig-2 double mutants, suggesting that other Rac GEFs may act in parallel to unc-73 (Shakir et al., 2006; Dyer et al., 2010). Recently, another DH domain protein-encoding gene, pix-1, was shown to display defects similar to unc-73 (Dyer et al., 2010). Double mutant analysis suggests that unc-73 and pix-1 function in parallel. Furthermore, pix-1 was shown to enhance the mig-2 but not the ced-10 phenotype, indicating that pix-1 and ced-10 function in a linear pathway. Based on these findings, a model has been proposed in which unc-73 activates both ced-10 and mig-2, while pix-1 activates ced-10 as part of a parallel pathway (Figure 3) (Dyer et al., 2010).
Importantly, unlike the genes described in Section 3.1, loss of genes involved in cytoskeletal remodeling do not affect the direction of Q cell migration.
After the initial migration, both Q neuroblasts divide and give rise to descendants that migrate further along the anteroposterior axis. Like the initial migration, Q descendant migration occurs in opposite directions, with the QL.x migrating towards the posterior and the QR.x towards the anterior (Figure 1, Figure 4, Movie 3, Movie 4). However, migration of the Q cell descendants depends on a different set of regulatory mechanisms and the initial left-right asymmetry components appear to be largely dispensable during this latter part of the Q cell migration process.
|
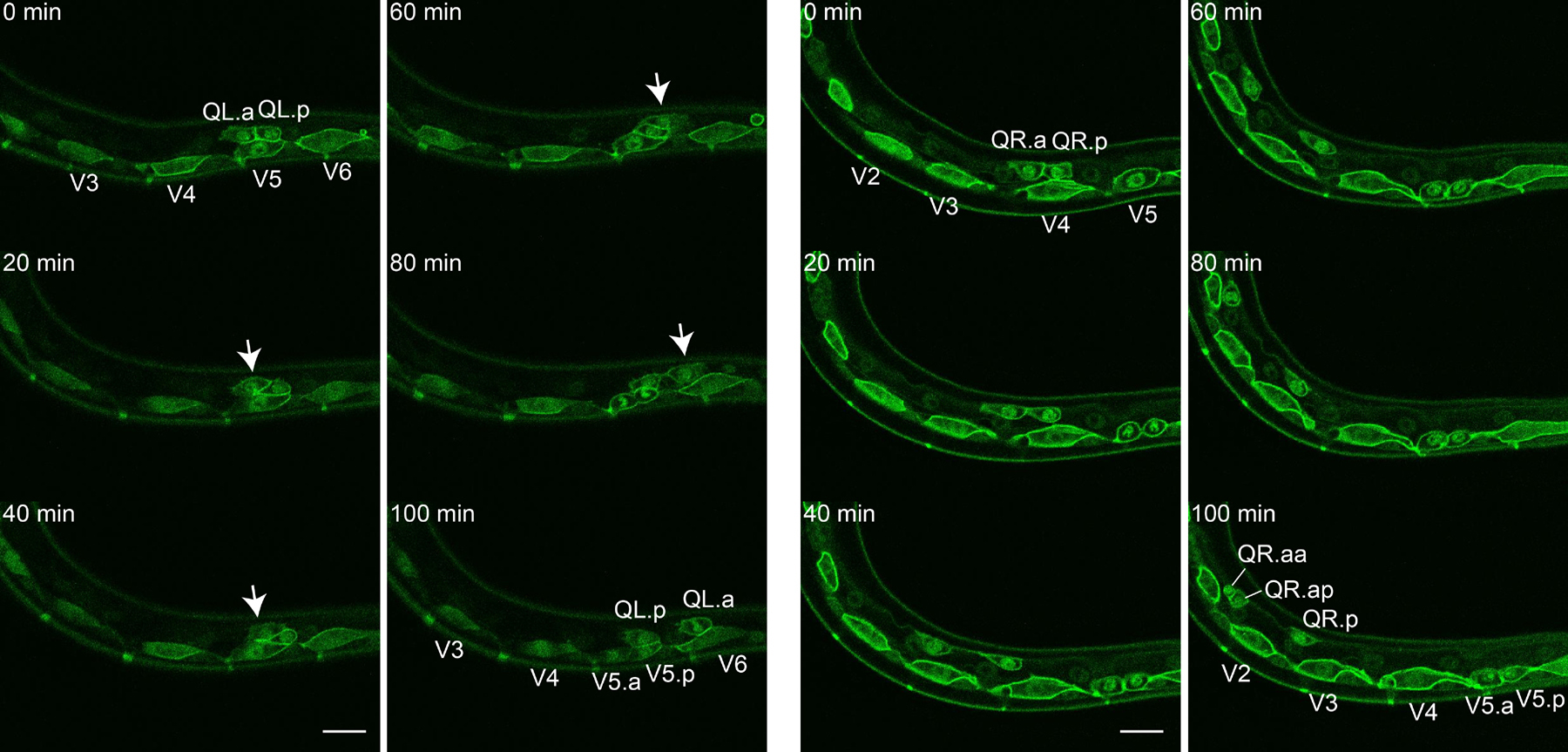
Figure 4. Q.x migration. Figure shows representative still images from Movie 3 (left two columns, QL.x) and Movie 4 (right two columns, QR.x). Arrows mark QL.a migrating over QL.p. At the 100 min time point QL.a displays a metaphase plate while QR.a has just divided in a small anterior daughter (QR.aa) and a larger posterior daughter (QR.ap). Scale bars depict 10 μm.
Movie 3: QL.x migration. Movie starts with QL.a and QL.p positioned above seam cell V5. QL.a migrates over QL.p towards the posterior. QL.p migrates a small distance and remains close to seam cell V5. In the final frame a metaphase plate appears in QL.a indicating the beginning of the second Q cell division. During the movie, seam cell V5 divides as well. Markers as in Movie 1. Ten min time intervals shown at 1 frame/s, dorsal is up, anterior is left.
Movie 4. QR.x migration. Movie shows QR.a and QR.p initially positioned on top of seam cell V4. Although both QR.a and QR.p migrate anteriorly, QR.a migrates faster (11.2 ± 2.6 μm/h) than QR.p (5.9 ± 2.2 μm/h) (Ou and Vale, 2009). Along its migratory path, QR.a executes the second Q cell division generating a small apoptotic anterior daughter cell (QR.aa) and a larger posterior daughter cell (QR.ap) that will migrate further towards the anterior. In the movie, seam cell V5 divides as well. Markers as in Movie 1. Ten min time intervals shown at 1 frame/s, dorsal is up, anterior is left.
As reviewed in the Wormbook chapter Wnt signaling in C. elegans, genetic analysis has shown that canonical Wnt/β-catenin signaling plays an important role in the control of QL.x migration. Briefly, during initial Q cell migration, canonical Wnt/β-catenin signaling is activated in QL but not in QR. This canonical Wnt signaling cascade, which includes EGL-20/Wnt, LIN-17/Fz, MIG-1/Fz, MIG-5/Dvl, PRY-1/Axin, AXL-1/Axin, GSK-3/GSK3β, BAR-1/β-catenin, and POP-1/Tcf ultimately results in expression of the Hox gene mab-5 in QL (Salser and Kenyon, 1992; Harris et al., 1996; Maloof et al., 1999; Whangbo and Kenyon, 1999; Korswagen et al., 2000; Herman, 2001; Korswagen et al., 2002; Walston et al., 2006; Oosterveen et al., 2007; Silhankova and Korswagen, 2007) (see also Wnt signaling in C. elegans for a more detailed overview in that organism). Expression of mab-5 was shown to be both necessary and sufficient for posterior migration of Q descendants. Apart from EGL-20, the other C. elegans Wnts appear to be dispensable for QL.x migration (Zinovyeva et al., 2008).
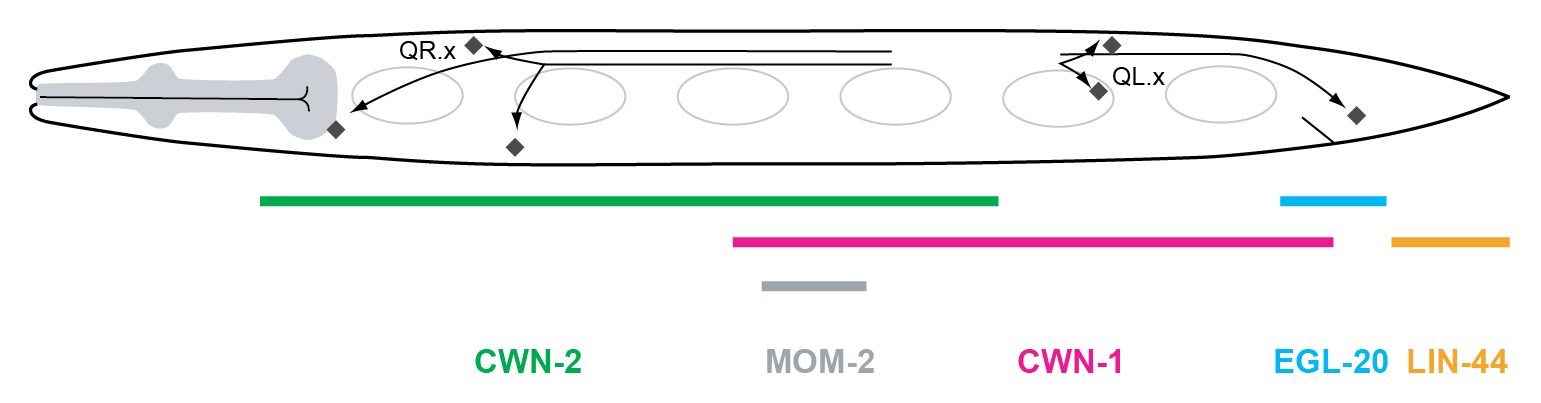
egl-20 is expressed in a group of cells in the tail region and exerts its function via long-range signaling. EGL-20 is distributed in an anteroposterior gradient, and formation of this gradient is necessary for QL to express mab-5 (Figure 5) (Whangbo and Kenyon, 1999; Coudreuse et al., 2006). As in other organisms, EGL-20 gradient formation requires a specialized Wnt secretion machinery, which includes the Wntless ortholog MIG-14 and the retromer complex (Banziger et al., 2006; Bartscherer et al., 2006; Coudreuse et al., 2006; Prasad and Clark, 2006; Harterink et al., 2011b).
|
Figure 5. Wnt gene expression during the first stage of larval development. The five C. elegans Wnt genes are expressed in partially overlapping domains along the anteroposterior axis of the L1 larva. lin-44 is expressed in the tail hypodermal cells hyp8-11, and in the rectal epithelial cells B and Y. egl-20 is expressed in the rectal epithelial cells K, F, U, B, Y, anal depressor muscle, P11/12, and in posterior ventral body wall muscle (BWM) quadrants. cwn-1 is expressed in posterior BWM, M cell descendants, anal depressor muscle, and CAN neurons. mom-2 is expressed in germ cell precursors Z2/Z3 and their descendants and a few unidentified cells in the tail. cwn-2 is expressed in head neurons, anterior BWM, anterior P.n cells, and in anterior intestinal cells. cwn-2 expression is highest around the terminal bulb of the pharynx (Gleason et al., 2006; Kennerdell et al., 2009; Song et al., 2010; Harterink et al., 2011).
The role of EGL-20 in Q cell migration is not restricted to the activation of mab-5 in QL. In egl-20 mutants, the final position of the QR.x is shifted posteriorly. QR.x migration was shown to be BAR-1/β-catenin independent, indicating that this function of EGL-20 is mediated via a non-canonical Wnt signaling pathway (Whangbo and Kenyon, 1999). Since EGL-20 is synthesized in the tail region and forms an anteroposterior gradient (Whangbo and Kenyon, 1999; Coudreuse et al., 2006), it is tempting to speculate that EGL-20 acts as a repellent that drives migration of the QR.x towards the anterior. However, an argument against this hypothesis is the observation that anterior expression of egl-20 partially rescues the QR.x migration phenotype of egl-20 mutants (Whangbo and Kenyon, 1999).
Another Wnt homolog, cwn-1, is also expressed in the tail region during late embryogenesis and early L1 development (Figure 5) (Gleason et al., 2006; Pan et al., 2006; Harterink et al., 2011a). Similar to loss of egl-20, cwn-1 mutants display a QR.x undermigration phenotype. Furthermore, loss of cwn-2, a Wnt gene that is expressed more anteriorly, also results in subtle undermigration of the QR.x (Figure 5) (Zinovyeva and Forrester, 2005; Gleason et al., 2006; Zinovyeva et al., 2008; Kennerdell et al., 2009; Song et al., 2010; Harterink et al., 2011a). cwn-2 enhances the cwn-1 and the egl-20 phenotype, while a cwn-1; egl-20 cwn-2 triple mutant shows the most severe QR.x undermigration phenotype (Zinovyeva et al., 2008). Together, these findings demonstrate that multiple Wnts function partially redundantly in controlling the anterior migration of the QR.x. The other C. elegans Wnt genes, mom-2 and lin-44, do not appear to be involved in the control of Q cell migration even though they are expressed during larval development (Figure 5) (Zinovyeva et al., 2008; Harterink et al., 2011a).
As expected, the C. elegans Frizzleds are essential for QR.x migration. However, when single mutants were examined, only mom-5/Fz displayed significant QR.x undermigration, while mutation of additional Frizzleds such as cfz-2 and lin-17 enhanced the phenotype (Zinovyeva et al., 2008). These results demonstrate that mom-5 is essential for proper QR.x migration, while the other Frizzled orthologs play a minor role. Frizzled quadruple mutants (in which all four Frizzled homologs are mutated) show a QR.x migration phenotype comparable to cwn-1; egl-20 cwn-2 triple mutants, supporting the notion that these Wnts primarily signal through Frizzled receptors to control QR.x migration (Zinovyeva et al., 2008). There is, however, also an important role for the C. elegans Ror ortholog CAM-1, with significant QR.x undermigration in cam-1 null mutants (Kim and Forrester, 2003).
As stated above, the initial left-right asymmetry of the Q neuroblasts is maintained during Q.x migration by two distinct Wnt signaling pathways: a canonical Wnt/β-catenin pathway that mediates posterior migration of the QL.x, and a β-catenin independent non-canonical Wnt pathway that mediates anterior migration of the QR.x. Whangbo and Kenyon have shown that the left-right asymmetry in this Wnt signaling response is based on a difference in threshold for canonical Wnt/β-catenin pathway activation (Whangbo and Kenyon, 1999). Thus, using heat-shock mediated expression of egl-20 in an egl-20 mutant background, it was found that at low EGL-20 ligand levels, only QL activates canonical Wnt signaling, while at high ligand levels QR also activates the canonical Wnt/β-catenin pathway.
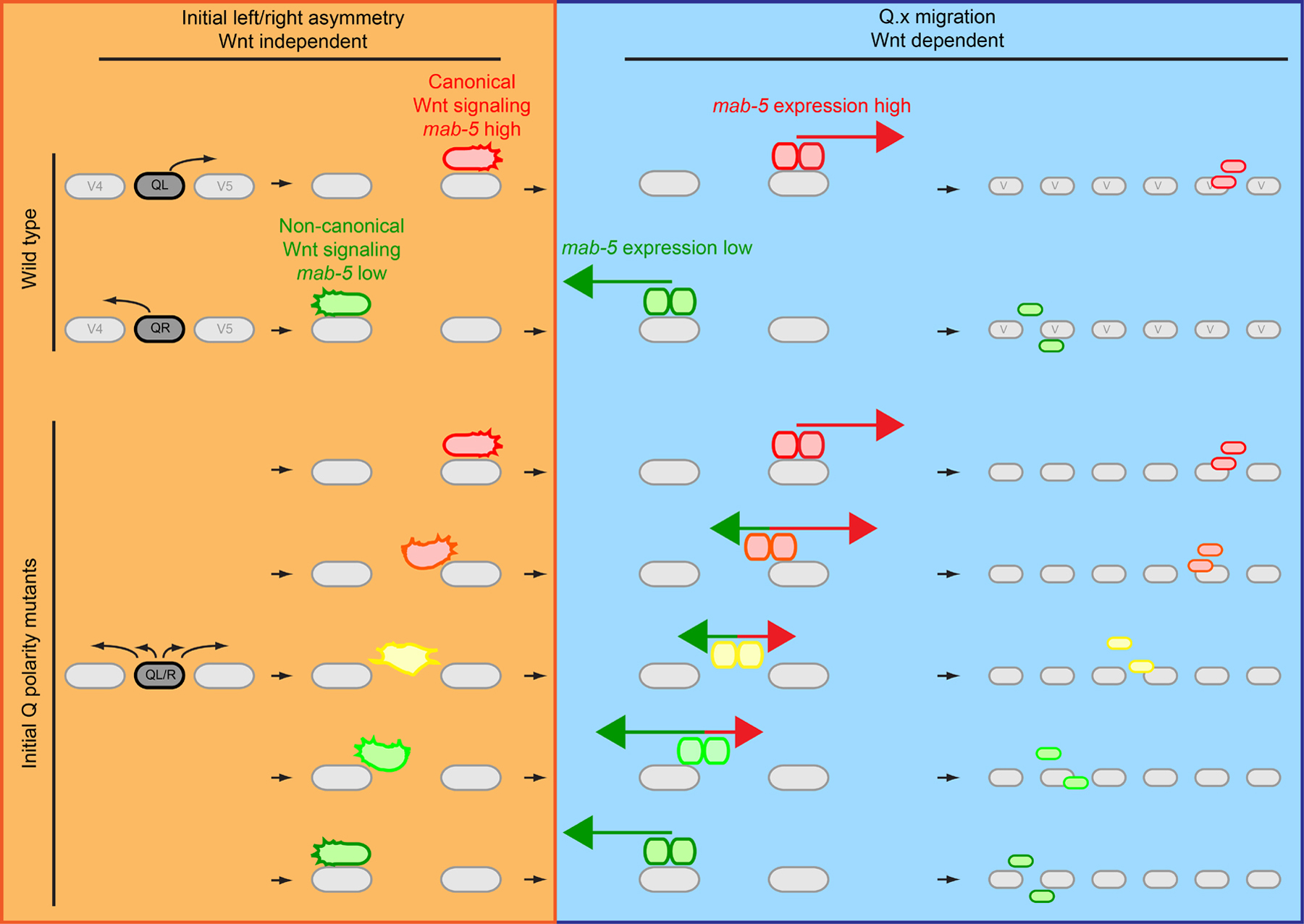
How is this difference in response threshold for canonical Wnt/β-catenin signaling determined? Interestingly, in mutants in which the initial left-right asymmetry of QL and QR polarization and migration is randomized (described in Section 3.1) the subsequent posterior and anterior migration of the QL.x and QR.x is also randomized (Honigberg and Kenyon, 2000; Du and Chalfie, 2001; Chapman et al., 2008; Middelkoop et al., 2012). Furthermore, the difference in activation threshold of QL and QR for canonical Wnt/β-catenin signaling is lost in mutants of the initial left-right asymmetry pathway (Middelkoop et al., 2012). These results suggest that the initial migration of the Q neuroblasts is mechanistically linked to the subsequent choice in Wnt signaling response.
Several lines of evidence suggest that the direction of initial Q cell polarization determines which Wnt signaling pathway is activated. Quantitative analysis of mab-5 expression (the target of Wnt/β-catenin signaling in the left Q neuroblast) using single molecule mRNA FISH (smFISH) in mutants that randomize the initial polarization of the Q neuroblasts revealed that posterior polarization correlates with high mab-5 expression, while anterior polarization correlates with low mab-5 expression. These results led to a model in which posterior polarization sensitizes the Q neuroblasts for canonical Wnt/β-catenin signaling, while anterior migration sensitizes for non-canonical Wnt signaling (Figure 6) (Honigberg and Kenyon, 2000; Du and Chalfie, 2001; Middelkoop et al., 2012). EGL-20 is present in a gradient that extends from the tail to the mid-body region of early L1 larvae (Whangbo and Kenyon, 1999; Coudreuse et al., 2006). Therefore, an interesting possibility is that posterior polarization of QL results in exposure to a higher level of EGL-20. In this model, the threshold for canonical Wnt pathway activation is not a Q cell intrinsic feature, but is dependent on the local concentration of EGL-20. Arguing against this model is the finding that reversal of the EGL-20 gradient rescues QL.x and QR.x migration in egl-20 mutants (Whangbo and Kenyon, 1999). Since initial polarization and migration is not affected in egl-20 mutants, these results suggest that the left-right asymmetry of Wnt signaling results from a Q cell intrinsic difference in Wnt sensitivity. An alternative model is that a local cue influences the Wnt signaling response of QL and QR. This model would also explain why the initial left-right asymmetric polarization and migration is instrumental for the left-right asymmetry in Wnt response.
|
Figure 6. Model of how the initial left-right asymmetry of the Q neuroblasts may dictate which Wnt signaling pathway is activated. Initial Q cell migration (top left side) is independent of Wnt signaling. In wild type, at the end of initial migration, canonical Wnt/β-catenin signaling is activated in QL resulting in expression of the Hox gene mab-5 (red), while QR activates non-canonical Wnt signaling and mab-5 expression is not upregulated (green). This is necessary for correct migration of the Q.x (right side), as canonical Wnt/β-catenin signaling drives posterior migration of the QL.x (red arrow) and non-canonical Wnt signaling drives anterior migration of the QR.x (green arrow, the panels on the right show the final positions of Q.paa and Q.pap). In mutants that affect the initial polarization of the Q neuroblasts, the direction of polarization and migration is random. In the bottom left side, different possible states of Q cell polarization in initial Q polarity mutants are depicted. In these mutants, the degree of posterior polarization correlates with activation of the canonical Wnt/β-catenin signaling pathway and high mab-5 expression (red). Anterior polarization correlates with activation of a non-canonical Wnt signaling response characterized by low mab-5 expression (green). Intermediate polarization states will result in intermediate levels of mab-5 expression (lower left panel, yellow). Depending on the degree of polarization, a tug of war between canonical and non-canonical Wnt signaling (middle right panels) may decide the direction and extent of Q.x migration.
The C. elegans genome contains six well-characterized Homeobox (Hox) genes that, like their metazoan and mammalian counterparts, are involved in anteroposterior patterning. During embryonic development, the Labial-like Hox gene ceh-13 is required for anterior specification, while the AbdB-like Hox genes nob-1 and php-3 specify posterior regions (Wittmann et al., 1997; Brunschwig et al., 1999; Van Auken et al., 2000). Three other Hox genes, lin-39, mab-5, and egl-5 are required for postembryonic patterning (Chisholm, 1991; Clark et al., 1993; Wang et al., 1993). In addition, ceh-13 was recently found to have a role in postembryonic development as well (Tihanyi et al., 2010).
As described in the previous section, the posteriorly expressed Hox gene mab-5 is a key regulator of QL.x migration (Salser and Kenyon, 1992). The proper migration of the Q descendants also requires lin-39. During L1 larval development, lin-39 is expressed in the mid-body region in ventral cord neurons, vulva precursor cells, and in Q neuroblasts and their descendants. While QL.x migration is largely unaffected in lin-39 mutants, the anterior migration of QR.x cells is aborted prematurely. Mosaic analysis indicates that lin-39 functions cell autonomously in this process (Clark et al., 1993; Wang et al., 1993). Unlike mab-5, lin-39 appears not to be regulated by EGL-20 signaling, as the QR.x undermigration phenotype of lin-39 and egl-20 single mutants is enhanced in the double mutant, indicating that they act in parallel pathways (Yang et al., 2005). However, it has not been tested whether this is also the case for cwn-1 and cwn-2.
Because of its critical function during embryonic patterning, the role of ceh-13 in postembryonic development has long been overlooked. However, Tihanyi and coworkers (2010) demonstrated a role for ceh-13 in Q.x migration and cell fusion during postembryonic development. Similar to lin-39, loss of ceh-13 results in premature termination of QR.x migration. In addition, ceh-13 induces a subtle but significant undermigration of the QL.x (Tihanyi et al., 2010).
Two Hox cofactors also play a role in Q.x migration. Mutations in the Hox cofactors ceh-20/Extradenticle and unc-62/Homothorax induce a clear undermigration of the QR.x. Furthermore, QL.ap is often positioned anterior to its wild type position in the tail region. Expression analysis of a ceh-20::gfp translational fusion revealed that ceh-20 is expressed in the Q neuroblasts and their descendants. Expression was also observed in P and V cells, possibly reflecting a role of ceh-20 in vulva development and V cell patterning, respectively. Furthermore, ceh-20::gfp expression was observed in all ventral cord neurons, several anterior neurons, body wall muscle cells, and in migratory cells like the mesoblast cell M and the BDU, ALM, and HSN neurons (Yang et al., 2005). The similarity in the QR.x migration phenotype of ceh-20 and unc-62 and the Hox genes lin-39 and ceh-13 suggests that unc-62 and ceh-20 may function as cofactors for these Hox genes. However, both unc-62 and ceh-20 enhance the QR.x migration phenotype of lin-39, indicating that they function (at least partially) independently of lin-39. Further analysis revealed that in lin-39 mutants, the QR.x are still susceptible to regulation by MIG-13/Lrp12 (see below) while in ceh-20 and unc-62 mutants they are not (Sym et al., 1999; Yang et al., 2005). These results suggest that ceh-20 and unc-62 are necessary for the QR.x to respond to the MIG-13 guidance cue.
mig-13/Lrp12 is a key regulator of QR.x migration (Harris et al., 1996; Sym et al., 1999). In mig-13 mutants, the QR.x are posteriorly displaced. Furthermore, mig-13 dramatically enhances the QR.x undermigration phenotype of egl-20 mutants. mig-13 encodes a single-pass transmembrane protein that is enriched on the cell surface. The predicted extracellular region has two conserved CUB domains, which mediate specific protein-protein interactions, and a single LDL receptor repeat (Bork and Beckmann, 1993; Sym et al., 1999). During corticogenesis of the mouse brain, the mig-13 homolog Lrp12 is expressed in a subpopulation of migrating neuronal cells in the preplate (Schneider et al., 2011). In addition, expression of mouse Lrp12 can partially rescue the QR.x migration defects of mig-13 mutants, consistent with the notion that mig-13 may have a conserved function in cell migration (Wang et al., 2013). To date, no interaction partners of MIG-13 have been identified.
Except for the posterior neuron DA9, mig-13 is predominantly expressed in the anterior half of the animal in the pharyngeal-intestinal valve cells and in neurons of the retrovesicular ganglion and the ventral cord. The expression of mig-13 is controlled by the Hox genes ceh-13 and mab-5, with ceh-13 activating mig-13 expression in the anterior and mab-5 inhibiting mig-13 expression in the posterior part of the body (Sym et al., 1999; Tihanyi et al., 2010). More recently, it was shown that mig-13 is also expressed in migrating QR.x cells (Wang et al., 2013). Interestingly, ubiquitous overexpression of mig-13 using a heat shock promoter revealed that MIG-13 promotes anterior migration in a dose-dependent manner, with overmigration of the QR.x at elevated doses of MIG-13 (Sym et al., 1999). Although initial studies using mosaic analysis suggested that mig-13 acts cell non-autonomously, a recent study has shown that Q neuroblast specific expression of mig-13 can rescue the mig-13 migration phenotype, indicating that mig-13 may have a cell autonomous function as well. Further studies are required to determine how MIG-13 controls Q neuroblast migration.
Like the initial migration of the Q neuroblasts, migration of the Q cell descendants is controlled by the partially redundantly acting Rho/Rac GTPases ced-10/Rac1 and mig-2/RhoG (for a detailed overview of small GTPases in C. elegans see Small GTPases). A mig-2 gain-of-function allele that encodes a constitutively active MIG-2 protein induces a widespread defect in cell migration (Zipkin et al., 1997). In these mig-2(gf) mutants, the Q.x migrate a shorter distance but the overall direction of migration is unaffected. Time-lapse analysis revealed that the onset and termination of Q.x migration as well as the timing of cell division is unaffected (Ou and Vale, 2009). However, the overall migration kinetics of the Q neuroblasts and their descendants are slower than in wild type (16.8 μm/h for QL.ap in wild type compared to 7.8 μm/h in mig-2 gain-of-function mutants). mig-2 loss-of-function results in similar but more subtle migration defects (Zipkin et al., 1997; Ou and Vale, 2009).
Strong mig-2 expression was observed in Q neuroblasts and their descendants, consistent with the idea that MIG-2 functions cell-autonomously (Zipkin et al., 1997). Furthermore, MIG-2 protein levels were found to be dynamically upregulated in cells with the highest migratory potential, such as QL.ap and QR.ap (Figure 1B) (Ou and Vale, 2009). This increase in MIG-2 protein levels may be a direct or indirect consequence of asymmetric segregation of cell fate determinants during the first Q neuroblast division.
Similar to what was described for the first phase of Q cell migration, the effect of mig-2 and ced-10 single mutants on Q.x migration is greatly enhanced in the double mutant. This enhancement was also observed when ced-10 or mig-2 loss-of-function mutations were combined with rac-2 RNAi. ced-10 is broadly expressed and its expression largely overlaps with that of mig-2 (Lundquist et al., 2001; Shakir et al., 2006). These findings demonstrate that the three C. elegans Rho/Rac orthologs function partially redundantly in the control of Q.x migration. Similar to the initial phase of Q cell migration, the guanine nucleotide exchange factors unc-73/Trio and pix-1 (Section 3.2) act in parallel to control Q.x migration, likely by activating the Rho/Rac GTPases (Figure 3) (Dyer et al., 2010).
In many migrating cells, Enabled/VASP proteins are highly enriched at filopodial tips and antagonize actin capping, a process that terminates actin filament nucleation (Welch and Mullins, 2002; Bear and Gertler, 2009). unc-34 encodes the C. elegans ortholog of Enabled/VASP and was shown to act in Q neuroblast migration (Yu et al., 2002; Withee et al., 2004; Shakir et al., 2006). UNC-34 contains an EVH1 domain that recognizes proline-rich motifs on target proteins, an SH3 binding domains, and an EVH2 domain thought to be necessary for tetramerization and actin binding. Truncation of a large part of the EVH2 domain yields a hypomorphic allele, indicating that UNC-34 tetramerizes and is involved in the regulation of actin dynamics (Bachmann et al., 1999; Zimmermann et al., 2002; Withee et al., 2004). unc-34 mutants show a subtle Q.x migration defect, in which Q descendants prematurely terminate their migration. Furthermore, unc-34 enhances the Q.x migration phenotype of rac-2 RNAi, ced-10, and mig-2 (Shakir et al., 2006). However, due to redundancy between the Rac1-like genes, it remains to be established whether unc-34 functions in a parallel pathway.
mig-15/NIK is also involved in the control of Q.x migration, in addition to its role in initial polarization and migration (see Section 3.1). The Q.x cells were found to migrate a shorter distance than in wild type animals, potentially reflecting a defect in the core cell migration machinery (Shakir et al., 2006; Chapman et al., 2008). Interestingly, loss of mig-2 enhanced the defect in Q.x migration direction found in weak alleles of mig-15. The phenotype of mig-15 mig-2 double mutants resembles that of stronger mig-15 alleles, indicating that mig-2 and mig-15 may function in the same pathway to control Q.x migration (Shakir et al., 2006).
To date, two cell adhesion proteins have been found to influence Q.x migration. First, loss of ina-1 (one of the two alpha-integrin subunits) was found to result in a reduction in the speed and extent of Q.x migration. Expression analysis revealed that INA-1::GFP is expressed in the Q.x. Furthermore, expression of ina-1 inversely correlates with migration speed (Baum and Garriga, 1997; Ou and Vale, 2009). These observations led to the hypothesis that low levels of integrin-based adhesion will result in high migratory speed, while high levels of integrin-based adhesion will reduce Q.x migration speed.
Second, it was demonstrated that the Fat-like cadherin CDH-4 controls the direction Q.x cell migration (Schmitz et al., 2008). Fat-like cadherins are characterized by a large number of extracellular cadherin repeats which are often accompanied by other sequence motifs like EGF repeats (Tanoue and Takeichi, 2005). cdh-4 encodes a Fat-like cadherin with 32 N-terminal cadherin domains and four EGF repeats that are separated by a single laminin G domain close to the membrane (Schmitz et al., 2008). Mutation of cdh-4 result in a variety of defects, including compromised axon guidance, aberrant hypodermal and pharyngeal development and Q.x cell migration defects. In these mutants, the QR.x often migrate posteriorly and the QL.x anteriorly. Furthermore, similar to initial Q polarity genes (see Section 3.1 and Section 3.3), cdh-4 appears to act upstream of mab-5. It is therefore likely that cdh-4 functions during the initial establishment of left-right asymmetry. Analysis of a cdh-4::gfp reporter gene revealed that cdh-4 is not expressed in Q neuroblasts, suggesting a cell non-autonomous role (Schmitz et al., 2008).
Over the years, studies on Q cell migration have unraveled many of the key mechanisms that control this highly stereotypic process. A complex network of Wnt signaling pathways, together with the action of homeotic genes, provides directionality and positions the Q descendants at precisely defined final positions along the anteroposterior axis. More downstream, a variety of cytoskeletal remodelers have been identified that mediate and control the motility of the Q neuroblasts. However, how positional information impinges on the core cell migration machinery remains to be established. Furthermore, migration potential appears to be hardwired into the Q cell lineage (Ou and Vale, 2009), but the mechanisms that specify the migration speed of different Q descendants are still largely unknown. The recently developed live imaging techniques that enable time-lapse imaging of Q neuroblast migration (Ou and Vale, 2009; Chai et al., 2012) provide a powerful tool to further dissect these mechanisms.
The left-right asymmetry of the initial migration of the Q cells is independent of Wnt signaling. However, Wnt signaling maintains the left-right asymmetry during Q descendant migration. So far, it has been shown that there is a clear correlation between initial posterior migration and activation of canonical Wnt/β-catenin signaling. However, the mechanistic link between these two processes remains largely unknown. The posteriorly polarized QL neuroblast may be exposed to a higher level of the EGL-20 ligand, which in turn triggers mab-5 expression. In this model, Q neuroblasts are not intrinsically different. However, several lines of evidence argue against this model. Alternatively, an as-of-yet unidentified external cue may modulate the response towards EGL-20. Future experiments may discriminate between these possibilities and thereby provide insight into how the initial left-right asymmetry is translated into Wnt signaling specificity.
Table 1. Genes involved in Q cell migration
| Gene | Q cell migration phenotype | Other comments |
|---|---|---|
| Initial polarization and migration: | ||
|
unc-40/DCC dpy-19/mDpy19 mig-15/NIK ptp-3/LAR |
Random polarization and migration of QL and QR | |
| Posterior migration of the QL.x: | ||
|
egl-20/Wnt lin-17/Fz mig-1/Fz bar-1/β-catenin pop-1/Tcf |
Anterior QL.x migration | Components of a canonical Wnt/β-catenin pathway that regulates mab-5/Hox expression |
|
pry-1/Axin axl-1/Axin gsk-3/GSK3β |
Posterior QR.x migration | Negative regulators of canonical Wnt signaling |
| Anterior migration of the QR.x: | ||
|
egl-20/Wnt cwn-1/Wnt cwn-2/Wnt mom-5/Fz cam-1/Ror |
Undermigration of the QR.x | Components of non-canonical Wnt signaling |
| Homeotic genes: | ||
|
mab-5/Antp lin-39/DfdScr ceh-13/Labial ceh-20/Exd unc-62/Meis |
Anterior QL.x migration QR.x undermigration |
mab-5 is necessary and sufficient for posterior Q.x migration |
| Cytoskeletal remodeling: | ||
|
mig-2/Mtl ced-10/Rac1 rac-2/Rac1 unc-73/Trio unc-34/Ena |
Q polarization defects and Q.x undermigration |
|
| Others: | ||
|
mig-13/Lrp12 ina-1/α-integrin cdh-4/Fat-like cadherin |
QR.x undermigration Q.x undermigration Loss of left/right asymmetry in Q.x migration |
Guidance cue controlled by mab-5 and ceh-13 |
We thank Hitoshi Sawa for critically reading the manuscript. This work was supported by a grant from the Dutch Cancer Society (HUBR 2008-4114) to H.C.K.
Bachmann, C., Fischer, L., Walter, U., and Reinhard, M. (1999). The EVH2 domain of the vasodilator-stimulated phosphoprotein mediates tetramerization, F-actin binding, and actin bundle formation. J. Biol. Chem. 274, 23549-23557. Abstract Article
Banziger, C., Soldini, D., Schutt, C., Zipperlen, P., Hausmann, G., and Basler, K. (2006). Wntless, a conserved membrane protein dedicated to the secretion of Wnt proteins from signaling cells. Cell 125, 509-522. Abstract Article
Bartscherer, K., Pelte, N., Ingelfinger, D., and Boutros, M. (2006). Secretion of Wnt ligands requires Evi, a conserved transmembrane protein. Cell 125, 523-533. Abstract Article
Baum, P.D., and Garriga, G. (1997). Neuronal migrations and axon fasciculation are disrupted in ina-1 integrin mutants. Neuron 19, 51-62. Abstract Article
Baumgartner, M., Sillman, A.L., Blackwood, E.M., Srivastava, J., Madson, N., Schilling, J.W., Wright, J.H., and Barber, D.L. (2006). The Nck-interacting kinase phosphorylates ERM proteins for formation of lamellipodium by growth factors. Proc. Natl. Acad. Sci. U. S. A. 103, 13391-13396. Abstract Article
Bear, J.E., and Gertler, F.B. (2009). Ena/VASP: towards resolving a pointed controversy at the barbed end. J. Cell Sci. 122, 1947-1953. Abstract
Bork, P., and Beckmann, G. (1993). The CUB domain: a widespread module in developmentally regulated proteins. J. Mol. Biol. 231, 539-545. Abstract Article
Brunschwig, K., Wittmann, C., Schnabel, R., Burglin, T.R., Tobler, H., and Muller, F. (1999). Anterior organization of the Caenorhabditis elegans embryo by the labial-like Hox gene ceh-13. Development 126, 1537-1546. Abstract
Buettner, F.F., Ashikov, A., Tiemann, B., Lehle, L., and Bakker, H. (2013). C. elegans DPY-19 is a C-mannosyltransferase glycosylating thrombospondin repeats. Mol. Cell 50, 295-302. Abstract Article
Carson, A.R., Cheung, J., and Scherer, S.W. (2006). Duplication and relocation of the functional DPY19L2 gene within low copy repeats. BMC Genomics 7, 45. Abstract Article
Ch'ng, Q., Williams, L., Lie, Y.S., Sym, M., Whangbo, J., and Kenyon, C. (2003). Identification of genes that regulate a left-right asymmetric neuronal migration in Caenorhabditis elegans. Genetics 164, 1355-1367. Abstract
Chai, Y., Li, W., Feng, G., Yang, Y., Wang, X., and Ou, G. (2012). Live imaging of cellular dynamics during Caenorhabditis elegans postembryonic development. Nature Prot. 7, 2090-2102. Abstract Article
Chalfie, M., and Sulston, J. (1981). Developmental genetics of the mechanosensory neurons of Caenorhabditis elegans. Dev. Biol. 82, 358-370. Abstract Article
Chalfie, M., Tu, Y., Euskirchen, G., Ward, W.W., and Prasher, D.C. (1994). Green fluorescent protein as a marker for gene expression. Science 263, 802-805. Abstract Article
Chan, S.S., Zheng, H., Su, M.W., Wilk, R., Killeen, M.T., Hedgecock, E.M., and Culotti, J.G. (1996). UNC-40, a C. elegans homolog of DCC (Deleted in Colorectal Cancer), is required in motile cells responding to UNC-6 netrin cues. Cell 87, 187-195. Abstract Article
Chapman, J.O., Li, H., and Lundquist, E.A. (2008). The MIG-15 NIK kinase acts cell-autonomously in neuroblast polarization and migration in C. elegans. Dev. Biol. 324, 245-257. Abstract Article
Chien, S.C., Brinkmann, E.M., Teuliere, J., and Garriga, G. (2013). Caenorhabditis elegans PIG-1/MELK acts in a conserved PAR-4/LKB1 polarity pathway to promote asymmetric neuroblast divisions. Genetics 193, 897-909. Abstract Article
Chisholm, A. (1991). Control of cell fate in the tail region of C. elegans by the gene egl-5. Development 111, 921-932. Abstract
Clark, S.G., Chisholm, A.D., and Horvitz, H.R. (1993). Control of cell fates in the central body region of C. elegans by the homeobox gene lin-39. Cell 74, 43-55. Abstract Article
Cordes, S., Frank, C.A., and Garriga, G. (2006). The C. elegans MELK ortholog PIG-1 regulates cell size asymmetry and daughter cell fate in asymmetric neuroblast divisions. Development 133, 2747-2756. Abstract
Coudreuse, D.Y., Roel, G., Betist, M.C., Destree, O., and Korswagen, H.C. (2006). Wnt gradient formation requires retromer function in Wnt-producing cells. Science 312, 921-924. Abstract Article
Drewes, G., and Nurse, P. (2003). The protein kinase kin1, the fission yeast orthologue of mammalian MARK/PAR-1, localises to new cell ends after mitosis and is important for bipolar growth. FEBS Latt. 554, 45-49. Abstract Article
Du, H., and Chalfie, M. (2001). Genes regulating touch cell development in Caenorhabditis elegans. Genetics 158, 197-207. Abstract
Dyer, J.O., Demarco, R.S., and Lundquist, E.A. (2010). Distinct roles of Rac GTPases and the UNC-73/Trio and PIX-1 Rac GTP exchange factors in neuroblast protrusion and migration in C. elegans. Small GTPases 1, 44-61. Abstract
Gleason, J.E., Szyleyko, E.A., and Eisenmann, D.M. (2006). Multiple redundant Wnt signaling components function in two processes during C. elegans vulval development. Dev. Biol. 298, 442-457. Abstract Article
Harbuz, R., Zouari, R., Pierre, V., Ben Khelifa, M., Kharouf, M., Coutton, C., Merdassi, G., Abada, F., Escoffier, J., Nikas, Y., et al. (2011). A recurrent deletion of DPY19L2 causes infertility in man by blocking sperm head elongation and acrosome formation. Am. J. Hum. Genet. 88, 351-361. Abstract Article
Harris, J., Honigberg, L., Robinson, N., and Kenyon, C. (1996). Neuronal cell migration in C. elegans: regulation of Hox gene expression and cell position. Development 122, 3117-3131. Abstract
Harterink, M., Kim, D.H., Middelkoop, T.C., Doan, T.D., van Oudenaarden, A., and Korswagen, H.C. (2011a). Neuroblast migration along the anteroposterior axis of C. elegans is controlled by opposing gradients of Wnts and a secreted Frizzled-related protein. Development 138, 2915-2924. Abstract Article
Harterink, M., Port, F., Lorenowicz, M.J., McGough, I.J., Silhankova, M., Betist, M.C., van Weering, J.R., van Heesbeen, R.G., Middelkoop, T.C., Basler, K., et al. (2011b). A SNX3-dependent retromer pathway mediates retrograde transport of the Wnt sorting receptor Wntless and is required for Wnt secretion. Nat. Cell Biol. 13, 914-923. Abstract Article
Hedgecock, E.M., Culotti, J.G., Hall, D.H., and Stern, B.D. (1987). Genetics of cell and axon migrations in Caenorhabditis elegans. Development 100, 365-382. Abstract
Herman, M. (2001). C. elegans POP-1/TCF functions in a canonical Wnt pathway that controls cell migration and in a noncanonical Wnt pathway that controls cell polarity. Development 128, 581-590. Abstract
Honigberg, L., and Kenyon, C. (2000). Establishment of left/right asymmetry in neuroblast migration by UNC-40/DCC, UNC-73/Trio and DPY-19 proteins in C. elegans. Development 127, 4655-4668. Abstract
Kennerdell, J.R., Fetter, R.D., and Bargmann, C.I. (2009). Wnt-Ror signaling to SIA and SIB neurons directs anterior axon guidance and nerve ring placement in C. elegans. Development 136, 3801-3810. Abstract Article
Kim, C., and Forrester, W.C. (2003). Functional analysis of the domains of the C. elegans Ror receptor tyrosine kinase CAM-1. Dev. Biol. 264, 376-390. Abstract Article
Kishi, M., Pan, Y.A., Crump, J.G., and Sanes, J.R. (2005). Mammalian SAD kinases are required for neuronal polarization. Science 307, 929-932. Abstract Article
Korswagen, H.C., Coudreuse, D.Y., Betist, M.C., van de Water, S., Zivkovic, D., and Clevers, H.C. (2002). The Axin-like protein PRY-1 is a negative regulator of a canonical Wnt pathway in C. elegans. Genes Dev. 16, 1291-1302. Abstract
Korswagen, H.C., Herman, M.A., and Clevers, H.C. (2000). Distinct β-catenins mediate adhesion and signalling functions in C. elegans. Nature 406, 527-532. Abstract Article
Kubiseski, T.J., Culotti, J., and Pawson, T. (2003). Functional analysis of the Caenorhabditis elegans UNC-73B PH domain demonstrates a role in activation of the Rac GTPase in vitro and axon guidance in vivo. Mol. Cell. Biol. 23, 6823-6835. Abstract
La Carbona, S., Allix, C., Philippe, M., and Le Goff, X. (2004). The protein kinase kin1 is required for cellular symmetry in fission yeast. Biol. Cell 96, 169-179. Abstract Article
Li, W., Zou, W., Yang, Y., Chai, Y., Chen, B., Cheng, S., Tian, D., Wang, X., Vale, R.D., and Ou, G. (2012). Autophagy genes function sequentially to promote apoptotic cell corpse degradation in the engulfing cell. J. Cell Biol. 197, 27-35. Abstract Article
Lizcano, J.M., Goransson, O., Toth, R., Deak, M., Morrice, N.A., Boudeau, J., Hawley, S.A., Udd, L., Makela, T.P., Hardie, D.G., et al. (2004). LKB1 is a master kinase that activates 13 kinases of the AMPK subfamily, including MARK/PAR-1. EMBO J. 23, 833-843. Abstract Article
Lundquist, E.A. (2006). Small GTPases. WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.67.1, http://www.wormbook.org.
Lundquist, E.A., Reddien, P.W., Hartwieg, E., Horvitz, H.R., and Bargmann, C.I. (2001). Three C. elegans Rac proteins and several alternative Rac regulators control axon guidance, cell migration and apoptotic cell phagocytosis. Development 128, 4475-4488. Abstract
Madaule, P., Furuyashiki, T., Eda, M., Bito, H., Ishizaki, T., and Narumiya, S. (2000). Citron, a Rho target that affects contractility during cytokinesis. Microsc. Res. Tech. 49, 123-126. Abstract Article
Mahmoudi, T., Li, V.S., Ng, S.S., Taouatas, N., Vries, R.G., Mohammed, S., Heck, A.J., and Clevers, H. (2009). The kinase TNIK is an essential activator of Wnt target genes. EMBO J. 28, 3329-3340. Abstract Article
Maloof, J.N., Whangbo, J., Harris, J.M., Jongeward, G.D., and Kenyon, C. (1999). A Wnt signaling pathway controls hox gene expression and neuroblast migration in C. elegans. Development 126, 37-49. Abstract
Middelkoop, T.C., Williams, L., Yang, P.T., Luchtenberg, J., Betist, M.C., Ji, N., van Oudenaarden, A., Kenyon, C., and Korswagen, H.C. (2012). The thrombospondin repeat containing protein MIG-21 controls a left-right asymmetric Wnt signaling response in migrating C. elegans neuroblasts. Dev. Biol. 361, 338-348. Abstract Article
Oosterveen, T., Coudreuse, D.Y., Yang, P.T., Fraser, E., Bergsma, J., Dale, T.C., and Korswagen, H.C. (2007). Two functionally distinct Axin-like proteins regulate canonical Wnt signaling in C. elegans. Dev. Biol. 308, 438-448. Abstract Article
Ou, G., Stuurman, N., D'Ambrosio, M., and Vale, R.D. (2010). Polarized myosin produces unequal-size daughters during asymmetric cell division. Science 330, 677-680. Abstract Article
Ou, G., and Vale, R.D. (2009). Molecular signatures of cell migration in C. elegans Q neuroblasts. J. Cell Biol. 185, 77-85. Abstract Article
Pan, C.L., Howell, J.E., Clark, S.G., Hilliard, M., Cordes, S., Bargmann, C.I., and Garriga, G. (2006). Multiple Wnts and frizzled receptors regulate anteriorly directed cell and growth cone migrations in Caenorhabditis elegans. Dev. Cell 10, 367-377. Abstract Article
Pierre, V., Martinez, G., Coutton, C., Delaroche, J., Yassine, S., Novella, C., Pernet-Gallay, K., Hennebicq, S., Ray, P.F., and Arnoult, C. (2012). Absence of Dpy19l2, a new inner nuclear membrane protein, causes globozoospermia in mice by preventing the anchoring of the acrosome to the nucleus. Development 139, 2955-2965. Abstract Article
Prasad, B.C., and Clark, S.G. (2006). Wnt signaling establishes anteroposterior neuronal polarity and requires retromer in C. elegans. Development 133, 1757-1766. Abstract Article
Salser, S.J., and Kenyon, C. (1992). Activation of a C. elegans Antennapedia homologue in migrating cells controls their direction of migration. Nature 355, 255-258. Abstract Article
Sawa H., and Korswagen H.C. Wnt signaling in C. elegans (December 9, 2013), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.7.2, http://www.wormbook.org.
Schmitz, C., Wacker, I., and Hutter, H. (2008). The Fat-like cadherin CDH-4 controls axon fasciculation, cell migration and hypodermis and pharynx development in Caenorhabditis elegans. Dev. Biol. 316, 249-259. Abstract Article
Schneider, S., Gulacsi, A., and Hatten, M.E. (2011). Lrp12/Mig13a reveals changing patterns of preplate neuronal polarity during corticogenesis that are absent in reeler mutant mice. Cereb. Cortex 21, 134-144. Abstract Article
Shakir, M.A., Gill, J.S., and Lundquist, E.A. (2006). Interactions of UNC-34 Enabled with Rac GTPases and the NIK kinase MIG-15 in Caenorhabditis elegans axon pathfinding and neuronal migration. Genetics 172, 893-913. Abstract Article
Silhankova, M., and Korswagen, H.C. (2007). Migration of neuronal cells along the anterior-posterior body axis of C. elegans: Wnts are in control. Curr. Opin. Genet. Dev. 17, 320-32. Abstract Article
Singhvi, A., Teuliere, J., Talavera, K., Cordes, S., Ou, G., Vale, R.D., Prasad, B.C., Clark, S.G., and Garriga, G. (2011). The Arf GAP CNT-2 regulates the apoptotic fate in C. elegans asymmetric neuroblast divisions. Curr. Biol. 21, 948-954. Abstract Article
Song, S., Zhang, B., Sun, H., Li, X., Xiang, Y., Liu, Z., Huang, X., and Ding, M. (2010). A Wnt-Frz/Ror-Dsh pathway regulates neurite outgrowth in Caenorhabditis elegans. PLoS Genet. 6 e1001056. Abstract Article
Steven, R., Kubiseski, T.J., Zheng, H., Kulkarni, S., Mancillas, J., Ruiz Morales, A., Hogue, C.W., Pawson, T., and Culotti, J. (1998). UNC-73 activates the Rac GTPase and is required for cell and growth cone migrations in C. elegans. Cell 92, 785-795. Abstract Article
Su, Y.C., Treisman, J.E., and Skolnik, E.Y. (1998). The Drosophila Ste20-related kinase misshapen is required for embryonic dorsal closure and acts through a JNK MAPK module on an evolutionarily conserved signaling pathway. Genes Dev. 12, 2371-2380. Abstract Article
Sulston, J.E., and Horvitz, H.R. (1977). Post-embryonic cell lineages of the nematode, Caenorhabditis elegans. Dev. Biol. 56, 110-156. Abstract Article
Sundararajan, L., and Lundquist, E.A. (2012). Transmembrane proteins UNC-40/DCC, PTP-3/LAR, and MIG-21 control anterior-posterior neuroblast migration with left-right functional asymmetry in Caenorhabditis elegans. Genetics 192, 1373-1388. Abstract Article
Sym, M., Robinson, N., and Kenyon, C. (1999). MIG-13 positions migrating cells along the anteroposterior body axis of C. elegans. Cell 98, 25-36. Abstract Article
Tanoue, T., and Takeichi, M. (2005). New insights into Fat cadherins. J. Cell Sci. 118, 2347-2353. Abstract Article
Tihanyi, B., Vellai, T., Regos, A., Ari, E., Muller, F., and Takacs-Vellai, K. (2010). The C. elegans Hox gene ceh-13 regulates cell migration and fusion in a non-colinear way. Implications for the early evolution of Hox clusters. BMC Dev. Biol. 10, 78. Abstract Article
Van Auken, K., Weaver, D.C., Edgar, L.G., and Wood, W.B. (2000). Caenorhabditis elegans embryonic axial patterning requires two recently discovered posterior-group Hox genes. Proc. Natl. Acad. Sci. U. S. A. 97, 4499-4503. Abstract Article
Wadsworth, W.G. (2002). Moving around in a worm: netrin UNC-6 and circumferential axon guidance in C. elegans. Trends Neurosci. 25, 423-429. Abstract Article
Walston, T., Guo, C., Proenca, R., Wu, M., Herman, M., Hardin, J., and Hedgecock, E. (2006). mig-5/Dsh controls cell fate determination and cell migration in C. elegans. Dev. Biol. 298, 485-497. Abstract Article
Wang, B.B., Muller-Immergluck, M.M., Austin, J., Robinson, N.T., Chisholm, A., and Kenyon, C. (1993). A homeotic gene cluster patterns the anteroposterior body axis of C. elegans. Cell 74, 29-42. Abstract Article
Wang, X., Zhou, F., Lv, S., Yi, P., Zhu, Z., Yang, Y., Feng, G., Li, W., and Ou, G. (2013). Transmembrane protein MIG-13 links the Wnt signaling and Hox genes to the cell polarity in neuronal migration. Proc. Natl. Acad. Sci. U.S.A. 110, 11175-11180. Abstract Article
Watanabe, K., Takebayashi, H., Bepari, A.K., Esumi, S., Yanagawa, Y., and Tamamaki, N. (2011). Dpy19l1, a multi-transmembrane protein, regulates the radial migration of glutamatergic neurons in the developing cerebral cortex. Development 138, 4979-4990. Abstract Article
Welch, M.D., and Mullins, R.D. (2002). Cellular control of actin nucleation. Annu. Rev. Cell Dev. Biol. 18, 247-288. Abstract Article
Whangbo, J., and Kenyon, C. (1999). A Wnt signaling system that specifies two patterns of cell migration in C. elegans. Mol. Cell 4, 851-858. Abstract Article
Wildwater, M., Sander, N., de Vreede, G., and van den Heuvel, S. (2011). Cell shape and Wnt signaling redundantly control the division axis of C. elegans epithelial stem cells. Development 138, 4375-4385. Abstract Article
Withee, J., Galligan, B., Hawkins, N., and Garriga, G. (2004). Caenorhabditis elegans WASP and Ena/VASP proteins play compensatory roles in morphogenesis and neuronal cell migration. Genetics 167, 1165-1176. Abstract Article
Wittmann, C., Bossinger, O., Goldstein, B., Fleischmann, M., Kohler, R., Brunschwig, K., Tobler, H., and Muller, F. (1997). The expression of the C. elegans labial-like Hox gene ceh-13 during early embryogenesis relies on cell fate and on anteroposterior cell polarity. Development 124, 4193-4200. Abstract
Wu, Y.C., Cheng, T.W., Lee, M.C., and Weng, N.Y. (2002). Distinct Rac activation pathways control Caenorhabditis elegans cell migration and axon outgrowth. Dev. Biol. 250, 145-155. Abstract Article
Xue, Y., Wang, X., Li, Z., Gotoh, N., Chapman, D., and Skolnik, E.Y. (2001). Mesodermal patterning defect in mice lacking the Ste20 NCK interacting kinase (NIK). Development 128, 1559-1572. Abstract
Yang, L., Sym, M., and Kenyon, C. (2005). The roles of two C. elegans HOX co-factor orthologs in cell migration and vulva development. Development 132, 1413-1428. Abstract Article
Yu, T.W., Hao, J.C., Lim, W., Tessier-Lavigne, M., and Bargmann, C.I. (2002). Shared receptors in axon guidance: SAX-3/Robo signals via UNC-34/Enabled and a Netrin-independent UNC-40/DCC function. Nat. Neurosci. 5, 1147-1154. Abstract Article
Zimmermann, J., Labudde, D., Jarchau, T., Walter, U., Oschkinat, H., and Ball, L.J. (2002). Relaxation, equilibrium oligomerization, and molecular symmetry of the VASP (336-380) EVH2 tetramer. Biochemistry 41, 11143-11151. Abstract Article
Zinovyeva, A.Y., and Forrester, W.C. (2005). The C. elegans Frizzled CFZ-2 is required for cell migration and interacts with multiple Wnt signaling pathways. Dev. Biol. 285, 447-461. Abstract Article
Zinovyeva, A.Y., Yamamoto, Y., Sawa, H., and Forrester, W.C. (2008). Complex network of Wnt signaling regulates neuronal migrations during Caenorhabditis elegans development. Genetics 179, 1357-1371. Abstract Article
Zipkin, I.D., Kindt, R.M., and Kenyon, C.J. (1997). Role of a new Rho family member in cell migration and axon guidance in C. elegans. Cell 90, 883-894. Abstract Article
Zou, W., Wang, X., Vale, R.D., and Ou, G. (2012). Autophagy genes promote apoptotic cell corpse clearance. Autophagy 8, 1267-1268. Abstract
*Edited by Joel H. Rothman and Andrew Singson Last revised September 18, 2013. Published October 15, 2014. This chapter should be cited as: Middelkoop, T.C., and Korswagen, H.C. Development and migration of the C. elegans Q neuroblasts and their descendants (October 15, 2014), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.173.1, http://www.wormbook.org.
Copyright: © 2014 Teije C. Middelkoop and Hendrik C. Korswagen. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
§To whom correspondence should be sent: Email: r.korswagen@hubrecht.eu
 All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.
All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.