Embryo series courtesy of Einhard Schierenberg
Embryo series courtesy of Einhard SchierenbergTable of Contents
Extracellular, intracellular, whole-cell and single-channel patch clamp recordings may all be made from the pharyngeal muscle. The type of recording made will be dictated by the information that is being sought. Extracellular recordings, termed electropharyngeograms or EPGs were pioneered by Leon Avery and co-workers (Avery et al., 1995) and provide information on parameters such as rate of pharyngeal pumping, duration of each pump (i.e. time between contraction and relaxation phases) and, by analysis of the shape of the EPG, the activity of neurones within the pharyngeal nervous system including the inhibitory neurone M3 and the pacemaker neurone MC (for details and examples see Raizen and Avery, 1994; Raizen et al., 1995). EPGs have the advantage of being relatively easy to record and yield information both on the activity of the muscle and the pharyngeal neurones. However, they give little reliable information on the magnitude of the pharyngeal potentials as the amplitude of the EPGs is very dependent on the positioning of the recording electrode and the properties of the seal with the worm's cuticle. In contrast, intracellular recordings are technically more challenging but are necessary if information on the resting membrane potential or shape of the pharyngeal action potential is required (e.g., Lee et al., 1997). Detailed analysis of the currents underlying the pharyngeal action potential may be obtained from whole-cell recording techniques. Although the pharyngeal muscle is not very amenable to voltage-clamp because of extensive electrical coupling between cells, nonetheless, it is possible to make a limited analysis of voltage-activated currents (Shtonda and Avery, 2005). Future progress on detailed analysis of the ionic currents in pharyngeal muscle may come from voltage-clamp studies on pharyngeal myoblasts in primary culture. Further information on the properties of single channels may also be obtained from patches excised from the pharynx, in either outside-out or inside-out configuration. The basic protocols for each of the electrophysiological techniques available for C. elegans pharynx are outlined below. For further background to the black art of microelectrode recording techniques consult Purves (1981).
A relatively stable recording table is required. An air-table is the best option but a stone slab, isolated from the frame and supported on rubber bungs, is usually quite sufficient provided that the environment is not mechanically very noisy.
An inverted microscope is better than an upright microscope as it provides more room to manoeuvre the electrode. The lamp should have an external DC power supply to limit problems with electrical noise. An X/Y moveable stage helps with positioning the preparation. Top of the range optics are not necessary, and x4 and x10 objectives will suffice.
A micromanipulator is needed to hold the suction electrode. As these are extracellular recordings, very fine movement is not required and a fairly coarse manual manipulator with fine movement in just the Z axis will suffice. Various options are available for mounting the manipulator on the recording stage, depending on the microscope being used, but the important consideration is stability. One option is to place the entire recording set-up on a metal surface (if using an air table this is the usual surface) and to use a manipulator with a magnetic base.
Electrical noise is one of the most severe limitations of the EPG technique as the amplitude of the EPG signal is relatively small. The apparatus should be enclosed in a Faraday cage to limit noise and to allow for earthing.
The recording chamber can be custom made from Perspex, or various options are available commercially. An easy and disposable alternative is to pour Sylgard at a depth of 0.5 cm in a 9 cm Petri dish. Allow it to set and then cut the required shape for the recording chamber out of the set Sylgard, allowing for a volume of approximately 1ml. Use a glass slide as the base and attach the Sylgard chamber to it. If the preparation is to be used for pharmacological experiments then important considerations are the volume of the bath versus the speed of the perfusion system to allow for rapid drug application and washing. To provide a stable reference potential, it is advisable to use an Agar bridge. This consists of a fine-bore glass tube filled with 4% Agar in 3 M KCl. The tube is bent so that one end can be placed in the recording chamber and the other in a small vial containing 3 M KCl to which the headstage ground is connected via a silver-chloride coated silver electrode.
A high-input impedance amplifier is required to record the EPGs, an analog to digital converter and a computer with software for data acquisition and to allow for off-line analysis.
A micropipette puller is required to pull the pipettes but a top of the range model is not necessary. A Narashige vertical drop puller for patch pipettes will suffice. The choice of pipette glass is probably not critical. Thick-walled borosilicate glass, 1 mm outer diameter without a filament is suitable. A pipette holder with a side-arm is used to hold the electrode.
All these items are obtainable from standard suppliers for biosciences research.
Pick animals into Dent's saline (composition in mM; D-glucose 10 mM, HEPES 10, NaCl 140, KCl 6, CaCl2 3, MgCl2 1, pH 7.4 with NaOH). It is important to consider the physiological status of the animals if comparisons are to be made between different groups. Therefore the age and nutritional status should be controlled for. Place the animals in a polystyrene tissue culture dish. The animals stick a little to the bottom of the dish which slows locomotion and makes it easier to cut the heads. An alternative method is to briefly chill the worms to immobilize them. Cut them (with a razor blade fragment held in a blade-holder) at the level of pharyngeal-intestinal valve. A solution of 0.1% bovine serum albumin (BSA) in Dent's saline should be used to release dissected tissue from the bottom of the Petri dish and ensure that it does not stick in pipettes. Transfer cut heads to the recording chamber in approx 5μl of 0.1% BSA Dent's using a Gilson pipette. To prepare the electrode for recording, pull a low resistance micropipette large enough to fit the head of the animal but small enough to provide a snug fit, i.e. about 10 μm diameter. A good pipette will last for half a dozen recordings or more if care is taken not to break it. Back-fill the micropipette with Dent's saline with a micropipette filler and attach it to the pipette holder. This has a silver-chloride coated silver wire to act as an electrode. The pipette holder should also have a side arm to which a length of tubing and a large (50 ml or more) syringe with a three-way tap is attached to enable suction to be applied. It is important that this is sealed well so that the suction cannot leak. Earth the apparatus to the signal ground input on the recording amplifier. Switch on the recording amplifier and set the tip potential to 0 mV using the voltage offset. Check the noise level. The EPGs are relatively small in amplitude and therefore the technique cannot tolerate a noise level of much more than 0.2 mV. Bring the electrode near to the anterior tip of the head and apply suction to the electrode thus attaching the head to the electrode. Close off the three-way tap to maintain suction. This helps to keep the head on the electrode. The noise will increase as the resistance of the seal increases. It may therefore be necessary to trouble-shoot additional sources of electrical noise (turning off the microscope lamp may help). If a perfusion system is being used then it can be started at this point. The amplitude and shape of the EPG are very dependent on the resistance of the seal and the relative placement of the electrode and therefore it is difficult to make comparisons between individual animals unless one is careful to control for these factors. Recordings may be made in voltage recording (‘Bridge’) mode. Recordings in either voltage-clamp or current-clamp do not add any advantage and simply introduce more electrical noise. This may necessitate filtering which can markedly distort the shape of the signal. Drugs may be applied via the perfusate or by aspiration of the bathing solution in the recording chamber.
EPG recordings are very stable and may be maintained for up to one hour or more, enabling detailed analysis of the response of the pharynx to different pharmacological agents. More care is required when making comparisons between mutant animals due to the intrinsic variability of the EPG shape. One aspect of EPG recordings that remains relatively limiting is that the analysis is still most accurately done manually, by cursor measurements on the digitized records. An example of an EPG is shown in Figure 1. There is scope for new development here in the form of software for reliable automated analysis of EPG shape.
|
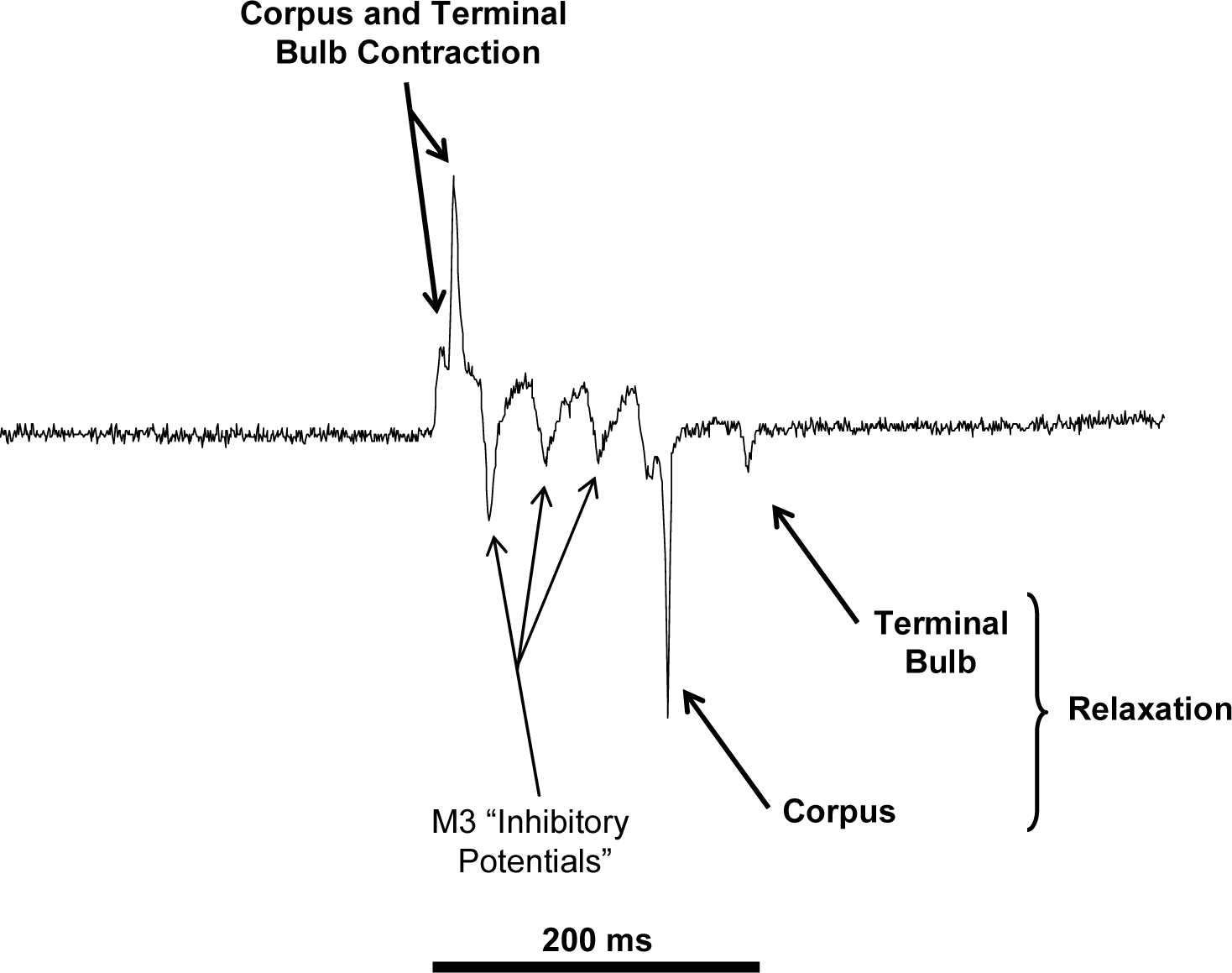
Figure 1. Example of an EPG recorded from a wild type (N2) pharynx. Note the separation of the positive going events corresponding to contraction of the corpus and terminal bulb, a number of “inhibitory potentials” correlating to activity of the M3 motorneurones. Negative going events are also to be observed which correspond to the relaxation of the corpus and terminal bulb respectively.
The equipment is similar to that required for EPG recordings except that some of the items are required to a higher specification. First, a very stable surface is required and it is advisable to use an anti-vibration table. Two manipulators are required, one coarse and one fine. These should be mounted on opposite sides of the microscope. The microscope will also require better optics than for EPG recordings e.g., a x40 objective. A recording amplifier such as the Axoclamp 2B (Axon Instrument, USA) or similar is required. The properties of the intracellular electrode are the key to making successful recordings from the pharynx. It must be robust enough to penetrate the tough basement membrane without damaging the small muscle cells. This requires a short shank with a very sharp tip. It is also advisable to use hard glass e.g., quartz or aluminosilicate (1mm outer diameter, with filament). Ideally, the electrodes should be pulled on a high specification horizontal puller (e.g., Sutter P2000 laser puller) capable of reproducibly pulling hard glass. The electrodes should be pulled to a resistance of 80 to 100 MΩ resistance, filled with 4M KAcetate; 10mM KCl.
Prepare cut heads and transfer them to the recording chamber as described for Protocol 1. Next pull a suction pipette, similar to, but with a slightly smaller tip opening than that used for the EPG recordings. The tip opening should fit over approximately one third of the terminal bulb. Place the suction electrode in an electrode holder (with a side-arm attached to a syringe via plastic tubing) and mount onto the coarse micromanipulator. Place the tip into the recording chamber and allow to back-fill with saline. Place the intracellular electrode in the pipette holder mounted in the fine manipulator on the opposite side of the microscope. This pipette will be used to hold the preparation in place and impart some structural rigidity to facilitate impalement of the terminal bulb muscle cell by the sharp electrode. Bring the tip of the suction pipette near to the terminal bulb of the pharynx and apply suction via the syringe so that the terminal bulb is held gently by the pipette against the bottom of the recording chamber. Then advance the intracellular recording electrode towards the opposite side of the terminal bulb until the tip dimples the muscle cell membrane without actually puncturing it. Intracellular access to the cell may be achieved by over-utilization of the capacitance compensation for the amplifier which causes the tip to cut through the membrane. (This ability is present on most amplifiers via a ‘buzz’ or ‘zap’ control). The resting membrane potential for wild type (N2) worms using standard Dent's saline outside is approximately −70 to −80mV. Every pharyngeal muscle pump coincides with an action potential that overshoots 0 mV by approximately 25 mV (Franks et al., 2002; Figure 2).
|
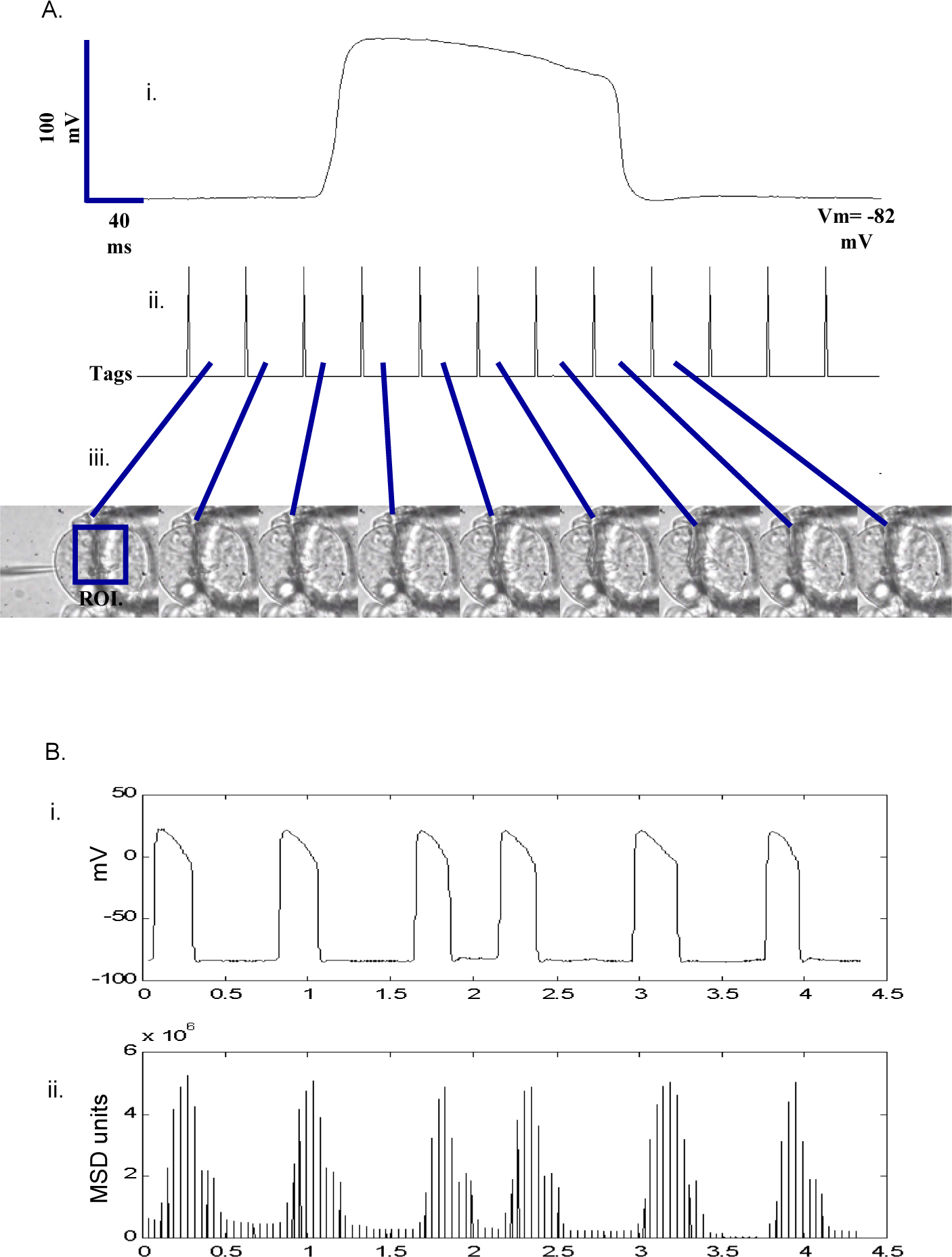
Figure 2. An automated system for the correlation of electrical potentials and muscle movement in the C. elegans pharynx. Intracellular recordings from the terminal bulb were made in the manner described. The muscle was simultaneously imaged using a black and white CCD. The CCD supplied a standard PAL, interlaced video signal from which the vertical sync was extracted using a simple circuit developed in-house. The extraction circuit was placed in-line between the video camera and a PC-based frame grabber. It consisted of an LM1881 chip, used to read vertical sync data and pass it to a monostable oscillator (a 555 chip) set to generate one TTL pulse (Tag) for every two fields in the video feed. In an interlaced video signal, two fields corresponds to one full frame of video. Therefore, every frame in the video sequence had an associated tag which was then passed to the Digidata acquisition system to be recorded simultaneously with the electrophysiological events. A single command switch simultaneously controlled the video feed to the frame grabber and also the output of the tag signal to ensure that the first and last recorded video frames corresponded to the first and final tags in the sequence sent to the data acquisition system. Each video frame was automatically analysed using a simple image/signal correlation algorithm. The degree of muscle movement between video frames was calculated and matched back to the timing tags in the electrophysiological recordings. Briefly, a region of interest ROI was defined manually and then signal intensity for each pixel in the ROI was automatically calculated. The mean square difference (MSD) of ROIs in adjacent frames was calculated and used as a measure of muscle movement. As the timing tags were based on the vertical sync signal of the PAL video feed MSD calculations were 40 ms apart. The figure shows output from the full recording setup. A. Each frame of the video (iii) signal can be directly associated with a TTL timing tag (ii) recorded simultaneously with the electrophysiological recording of membrane potential (i). The lumen of the terminal bulb can be seen to gradually open and close in each successive frame during the action potential. One action potential corresponds to one motor cycle in the terminal bulb. B. In this recording each full action potential (i) is reflected in the MSD trace (ii). The mean latency of peak muscle contraction to peak action potential amplitude in this experiment was approximately 150 ms primarily reflecting the latency associated with excitation-contraction coupling in the pharyngeal muscle. Only six action potentials are shown in the trace, which spans 4.5 seconds. However, the number of action potentials that can be recorded with corresponding video and then processed, is limited only by the hard disk capacity and processing power of the PC used.
As for Protocol 2. Although a voltage-clamp amplifier such as the Axoclamp 2B may be used for these studies, a patch-clamp amplifier such as the Axopatch 200B is preferable.
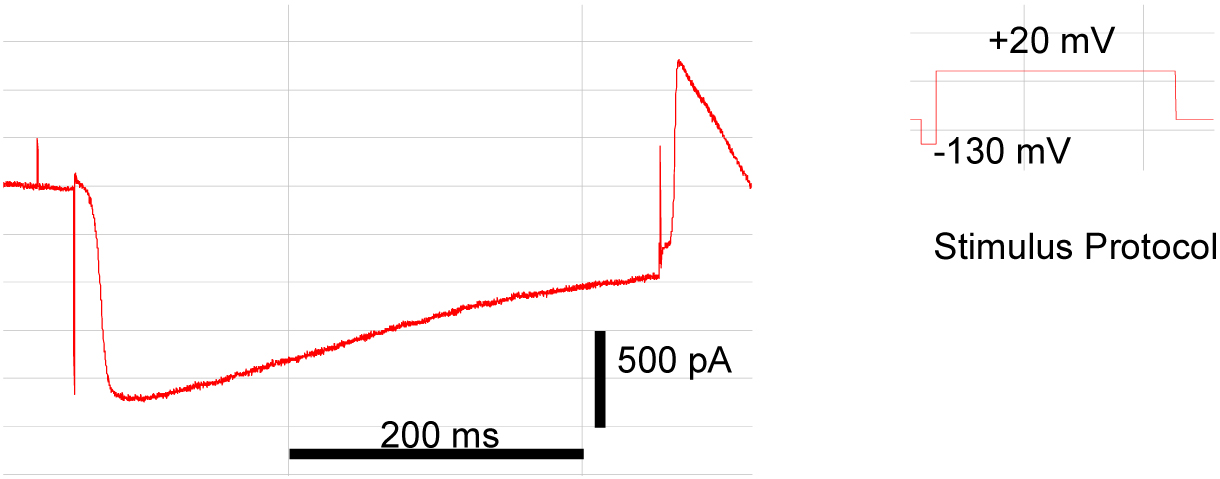
A protocol for recording whole cell currents from the corpus region of the pharyngeal muscle has been described by Avery (Shtonda and Avery, 2005). Recordings may also be made from the terminal bulb region (Vinogradova et al., 2006). The methods are similar to patch clamp techniques described for other cells with the additional challenge, and limitation, that the geometry of the pharyngeal muscle makes it less amenable to this approach and voltage control will not be accurate. Accordingly, care must be taken not to over interpret data from such studies. For recordings from the terminal bulb, heads are transferred to the recording chamber, which contains a suitable external saline e.g., Dent's saline. A suction pipette (filled with Dent's saline) is attached to the cut edge of the cuticle and used to gently hold the preparation against the base of the recording chamber. The pharynx of C. elegans is surrounded by a basement membrane which will interfere with seal formation and obtaining whole cell access. Therefore, before one can begin to record from the muscle cells one needs remove the basement membrane by enzymatic digestion. A method modified from (Richmond and Jorgensen, 1999) is suitable. First, treat the preparation with a solution of 0.1% trypsin in Dent's saline for 30 seconds and then wash gently with Dent's saline to remove trypsin. Next, treat for 2–4 minutes with a solution containing (in Dent's): 0.23 mg ml-1 Protease Type XIV and 0.62 mg ml-1 Collagenase P. (The exact timing of this second treatment is crucial for success and will have to be determined by the operator). Finally, wash gently with Dent's to stop digestion. Patch electrodes for whole-cell recordings may be pulled from Haematokrit capillaries (resistance 3–5 MΩ) and filled with appropriate internal solutions (e.g., composition, in mM: 150 N-methyl-D-glucamine, 4 MgCl2, 5 HEPES, 0.25 CaCl2, 36 sucrose, 5 EGTA; pH 7.2). Single-channel recordings may be performed in outside-out and inside-out configuration using thick-walled electrodes (GC150-7.5 glass, Harvard Apparatus). The electrode tips need to be coated with beeswax (to reduce electrode capacitance) and fire-polished. Using these methods seal resistances in the range of 10–30 GΩ can be achieved. Data may be acquired with a patch-clamp amplifier in the standard manner. However, a special consideration for whole cell recordings from pharynx is that the input capacitance is very large and cannot be reliably and accurately compensated. Doubtless this is because of the highly folded surface of the muscle cell membrane and extensive electrical coupling between cells. An example of a whole cell recording is shown in Figure 3.
Irina Vinogradova contributed to the development of the patch clamp protocols. Enriq Claverol and Christopher Franks developed the automated system for simultaneous acquisition of electrical and video signals. We gratefully acknowledge the pioneering work of the Avery lab who led the way in this field. The Holden-Dye group also acknowledge the support of the Biotechnology and Biological Science Research Council, UK for funding the electrophysiological analysis of C. elegans.
Avery, L., Raizen, D., and Lockery, S. (1995). Electrophysiological methods. Methods Cell Biol. 48, 251–269. Abstract
Franks, C.J., Pemberton, D., Vinogradova, I., Cook, A., Walker, R.J., and Holden-Dye, L. (2002). The ionic basis of the resting membrane potential and action potential in the pharyngeal muscle of Caenorhabditis elegans. J. Neurophysiol. 87, 954–961. Abstract
Lee, R.N., Lobel, L., Hengartner, M., Horvitz, H.R., and Avery, Y.L. (1997). Mutations in the alpha 1 subunit of an L-type voltage-activated Ca2+ channel cause myotonia in Caenorhabditis elegans. EMBO J. 16, 6066–6076. Abstract Article
Purves, R.D. (1981). Microelectrode Methods for Intracellular Recording and Ionophoresis (Academic Press).
Raizen, D.M., and Avery, L. (1994). Electrical-activity and behavior in the pharynx of Caenorhabditis elegans. Neuron 12, 483–495. Abstract Article
Raizen, D.M., Lee, R.N., and Avery, L. (1995). Interacting genes required for pharyngeal excitation by motor-neuron MC in Caenorhabditis elegans. Genetics 141, 1365–1382. Abstract
Richmond, J., and Jorgensen, E.M. (1999). One GABA and two acetylcholine receptors function at the C. elegans neuromuscular junction. Nat. Neurosci. 2, 791–797. Abstract Article
*Edited by William J. Schafer. WormMethods editor, Victor Ambros. Last revised August 10, 2005. Published May 17, 2006. This chapter should be cited as: Cook, A., et al. Electrophysiological recordings from the pharynx (May 17, 2006), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.110.1, http://www.wormbook.org.
Copyright: © 2006 Alan Cook, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
§To whom correspondence should be addressed. E-mail: lmhd@soton.ac.uk
 All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.
All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.