Embryo series courtesy of Einhard Schierenberg
Embryo series courtesy of Einhard SchierenbergTable of Contents
Abstract
Nuclear receptors (NRs) are transcription factors typically regulated by lipophilic hormones, which coordinate metazoan metabolism, development and homeostasis. C. elegans has undergone a remarkable expansion of the family, harboring 284 of these receptors in its genome. Approximately 20 of them have been analyzed genetically, most of which correspond to conserved homologs in other metazoans. These NRs variously affect broad life history traits such as sex determination, molting, developmental timing, diapause, and life span. They also impact neural development, axon outgrowth and neuronal identity. Finally, they influence lipid and xenobiotic metabolism. The study of C. elegans NRs holds great promise for dissecting nuclear receptor signaling pathways in vivo in the context of complex endocrine networks.
An early invention of metazoan signal transduction, nuclear receptors (NRs) comprise a family of transcription factors often regulated by small lipophilic molecules, such as steroids, retinoids, bile and fatty acids, that mediate endocrine control (Mangelsdorf et al., 1995). In addition, many so-called orphan NRs have either unidentified cognate ligands or none at all (Mangelsdorf and Evans, 1995). Although the vertebrate NRs are well-studied mechanistically, an exploration of NR function in a simple well-defined genetic organism such as C. elegans has illuminated their in vivo physiology, and their place in global regulatory networks.
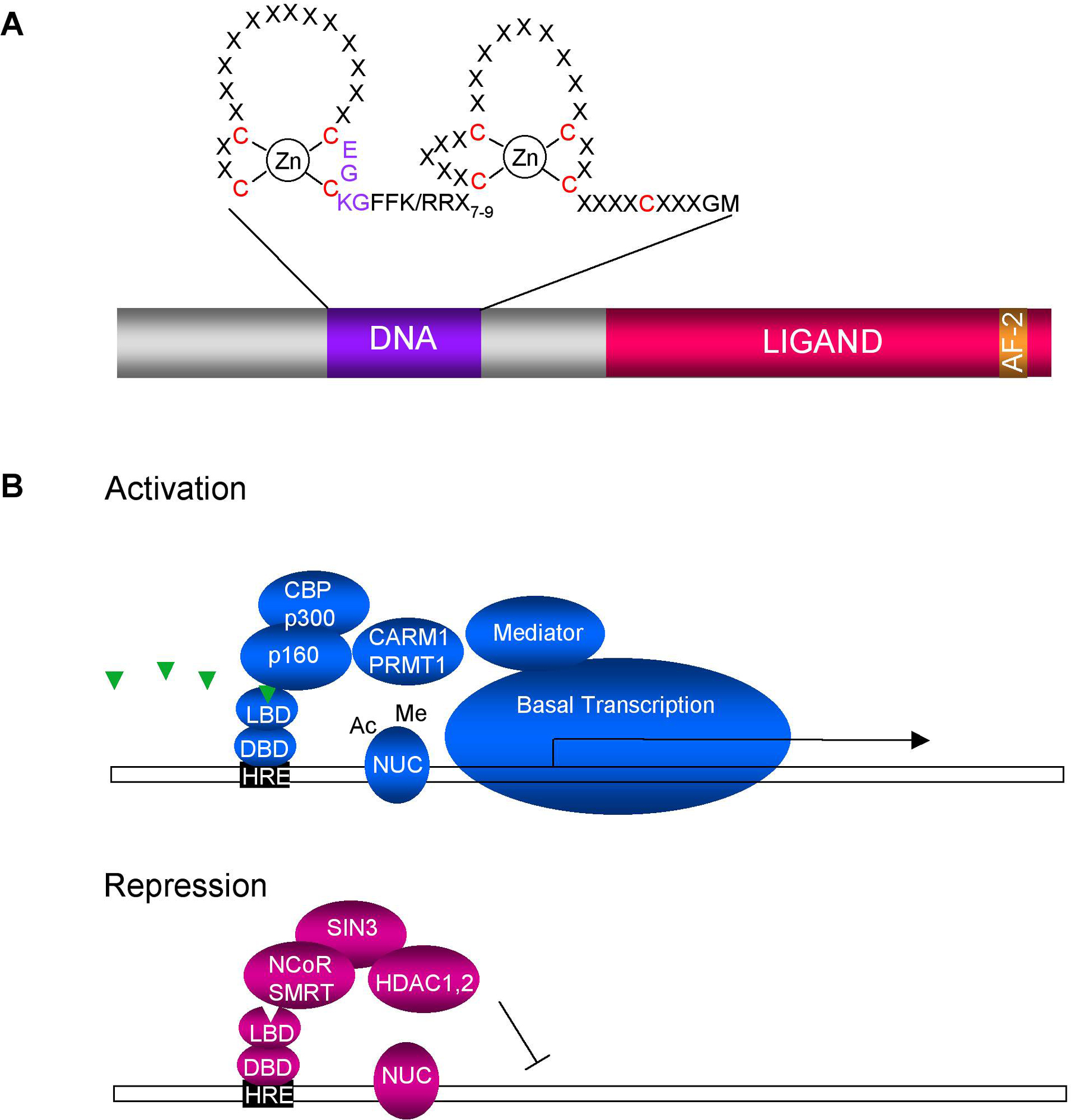
The key to NR signaling lies in their conserved molecular architecture (Mangelsdorf et al., 1995). The N-terminus contains a DNA binding domain (DBD) consisting of two Cys4 zinc fingers (Figure 1A). Much of the signaling intelligence resides in a C-terminal ligand binding domain (LBD), which not only sequesters ligand but also docks coactivators and corepressors—adaptor proteins that couple the NR to histone acetyltransferase and deacetylase complexes, respectively (Figure 1B). In addition, the LBD largely mediates receptor hetero- or homodimerization. Crystal structure and biochemical studies reveal that ligand binding triggers a conformational change in which a C-terminal transactivation helix, called AF-2, snaps back onto the LBD core (Bourguet et al., 1995; Renaud et al., 1995). Consequently, bound corepressors are displaced by coactivators, leading to gene expression. Moreover, NR activity is often regulated by phosphorylation, acetylation, and other covalent modifications (Fu et al., 2004).
|
Figure 1. (A) Schematic diagram of an exemplary nuclear hormone receptor. DNA binding domain (DNA) comprised of two Cys4 Zn fingers. Nine conserved cysteines are highlighted in red. A typical P-box, (CEGCKG) which confers DNA binding specificity, is shown. The ligand binding domain (ligand) contains multiple surfaces that recruit coactivators or corepressors. The AF-2 transactivation helix closes the ligand binding pocket during hormonal activation. (B) General model for transcriptional activation and repression based on vertebrate receptors. In the presence of hormone (triangle), NRs bind to their hormone response elements (HRE), and assemble coactivator complexes that acetylate (ac, p160, p300) or methylate (me, CARM1, PRMT1) nucleosomes (NUC). Mediator components contact NRs and facilitate the recruitment of the basal transcription machinery, turning on gene expression. Some nuclear receptors repress in the absence of hormone, by assembling corepressor complexes, such as SMRT and NCoR, which recruit histone deacetylases (HDACs) that inhibit transcription. The composition of activation and repression complexes depends on the NR as well as the promoter context. In addition, many NRs work as homo- or heterodimers.
Evidently, NRs have undergone an explosive expansion and divergence in the worm. C. elegans has an astounding 284 receptors, compared to 48 for humans and 21 for flies. (For comparative phylogenetic trees see Maglich et al., 2001; Robinson-Rechavi et al., 2005; Sluder and Maina, 2001; Sluder et al., 1999). Fifteen NRs have clear homologs in other species (Table 1), and include relatives of the mammalian HNF4, Vitamin D receptor, COUP-TF, SF1, ROR, PNR, GCNF, TLX as well as the Drosophila DHR3, DHR38, E75, E78, DHR78, and DHR96. The other 279 receptors arose from an ancestral HNF4 (Robinson-Rechavi et al., 2005). Why do worms have so many receptors? One thought is that gene duplication has been deployed instead of receptor combinatorics. Consistent with this, C. elegans lacks an apparent RXR/USP, a heterodimeric partner with numerous NRs (Sluder and Maina, 2001). However, yeast two-hybrid screens identify several interacting NRs (Li et al., 2004), suggesting C. elegans may have evolved a combinatorial system different from RXR.
Table 1. Some functionally described C. elegans NRs and their homologs
| C. elegans | Function | D. melanoga- ster | Function | M. musculus/ H. sapiens | Function |
|---|---|---|---|---|---|
| SEX-1 | Sex determination | E78 | Molting | revERB | Circadian clock, transcriptional repression |
| NHR-85 | Egg laying, molting? | E75 | Molting | ||
| NHR-23 | Molting, epidermal differentiation, dauer formation | DHR3 | Embryonic development, molting, metamorphosis | ROR alpha | Cerebellar differentiation |
| beta | Circadian clock | ||||
| gamma | Thymopoesis | ||||
| NHR-25 | Ventral closure, epidermal differentiation, molting, dauer formation | FTZ-F1 alpha | Segmentation | SF1 | Steroidogenesis |
| FTZ-F1beta | Molting, metamorphosis | LHR | Cholesterol homeostasis, bile acid metabolism | ||
| NHR-41 | Dauer formation | DHR78 | Molting, metamorphosis | TR2/TR4 | Unknown |
| NHR-6 | Ovulation | DHR38 | Molting | Nurr1 | Dopaminergic differentiation |
| NGFI-B/nur 77 | Apoptosis, immediate early response | ||||
| NHR-67 | Molting, growth, vulval formation | Tailless | A/P patterning, neurogenesis | TLX | Forebrain development, neural stem cell maintenance |
| NHR-91 | No obvious function | DHR4 | Unknown | GCNF | Germ cell differentiation, embryogenesis |
| DAF-12 | Dauer formation, lipid metabolism, stage specification, life span | DHR96 | Unknown | VitD | Bone differentiation, bile acid metabolism |
| CAR, PXR | Xenobiotic and bile metabolism | ||||
| NHR-8 | Xenobiotic metabolism | ||||
| NHR-48 | Unknown | ||||
| NHR-49 | Fatty acid metabolism | dmHNF4 | Unknown | HNF4 | Glucose homeostasis, liver metabolism |
| UNC-55 | Neural differentiation | Seven-up | Photoreceptor fate | COUP TF1 COUP TF2 | Neural development |
| FAX-1 | Neural differentiation | dmFAX-1 | Unknown | PNR | Photoreceptor fate |
| ODR-7 | Olfaction | None | None |
Only about twenty C. elegans NRs have described visible phenotypes. Below we highlight a handful.
Acting early in development, SEX-1 regulates C. elegans sex determination and dosage compensation by downregulating a sex determining gene called xol-1(Carmi et al., 1998). Encoded on the X chromosome, sex-1´s dose determines the level of xol-1 repression. When SEX-1 is high, XOL-1 is low, and animals develop as hermaphrodites. Conversely, when SEX-1 is low, XOL-1 is high, and animals become males. SEX-1 is most homologous to E78A, a Drosophila ecdysone-induced NR of unknown function (Stone and Thummel, 1993), and to vertebrate rev-Erb, an orphan NR that behaves as a transcriptional repressor in circadian oscillators (Preitner et al., 2002). The SEX-1 ligand binding domain is quite diverged, and it is unknown whether its activity is hormone regulated. Interestingly, C. elegans sex is also influenced by diet; paternal X disjunction is increased by unknown metabolites from log phase E. coli (Prahlad et al., 2003). Conceivably, such metabolites might somehow impinge on SEX-1.
Nematodes are postulated to belong to the Ecdysozoa, a proposed broad clade of animals that molt (Aguinaldo et al., 1997). In Drosophila and other insects, pulses of the hormone 20-hydroxyecdysone stimulate the ecdysone receptor, which initiates transcriptional cascades that drive molting and metamorphosis (Riddiford et al., 2003). Downstream, several NR transciption factors, including DHR3, FTZ-F1, E75/E78, DHR38, DHR78 and others are turned on in a stereotypical sequence (Ashburner, 1974; Riddiford et al., 2000; Sullivan and Thummel, 2003). Remarkably, C. elegans lacks ecdysone, the ecdysone receptor, and its heterodimeric partner USP. Little is known about how the molt cycle is driven. Conceivably, another sterol hormone does the job, since cholesterol deprivation, as well as disruption of genes implicated in sterol transport result in molting, growth or fecundity defects (Matyash et al., 2004; Merris et al., 2003; Shibata et al., 2003; Shim et al., 2002; Yochem et al., 1999). In addition, C. elegans harbors five orthologs of the ecdysone inducible NRs mentioned above. Several are expressed periodically, attuned to the molt cycle, and some mediate ecdysis although others do not (Asahina et al., 2000; Gissendanner et al., 2004; Gissendanner and Sluder, 2000; Kostrouchova et al., 2001).
C. elegans NHR-23 is a homolog of Drosophila DHR3, which mediates the pre-pupal to pupal transition in fly (Lam et al., 1999). Similarly, NHR-23 functions in ecdysis; RNAi knockdown results in molting defects at all four molts. Other phenotypes indicate aberrant epidermal differentiation, including disrupted collagen synthesis, epidermal seam cell displacement and blunted male tail development (Kostrouchova et al., 1998; Kostrouchova et al., 2001). Accordingly, NHR-23 is expressed in the epidermis. The vertebrate homologs, RORα,β,γ, function in various processes including Purkinje cell generation, circadian rhythms, and thymopoesis (Jetten et al., 2001). It may be significant that orthologs in all three species are part of biological clocks—the molt cycle and the circadian oscillator.
C. elegans NHR-25 belongs to a highly conserved receptor subtype that includes Drosophila FTZ-F1 and the human SF1 and LRH. Drosophila FTZ-F1 functions in embryonic segmentation and larval metamorphosis (Broadus et al., 1999; Guichet et al., 1997; Lavorgna et al., 1993; Ueda et al., 1990; Yu et al., 1997). Similarly, nhr-25 mutants arrest at the two-fold stage of embryogenesis, prior to elongation, with defects in ventral closure of the epidermis (Chen et al., 2004; Silhankova et al., 2005). Larvae also have defects in molting, epidermal and vulval cell fusion, and cell elongation. Adults exhibit aberrant somatic gonadal development, tumorous germlines, and are sterile (Asahina et al., 2000; Gissendanner et al., 2004; Gissendanner and Sluder, 2000; Hwang and Sternberg, 2004). Consistent with its phenotype, NHR-25 is expressed in epidermis and somatic gonad (Asahina et al., 2000; Gissendanner and Sluder, 2000). Interestingly, mammalian SF1 also controls differentiation of the gonad, as well as the adrenal gland, pituitary and hypothalamus, where it initiates transcription of key genes involved in steroidogenesis (Parker et al., 2002). LRH regulates bile acid and cholesterol metabolism (Fayard et al., 2004).
The rudimentary sketches of an ecdysis signaling pathway are beginning to take shape. Interestingly, let-767 encodes a 17-betahydroxysteroid reductase, which when mutated gives rise to molting defects and embryonic lethality reminiscent of NHR-25 (Kuervers et al., 2003). Conceivably, this protein works upstream in the production of a molting hormone. Downstream of both NHR-25 and NHR-23, acn-1 encodes a metalloprotease implicated in molting (Brooks et al., 2003).Work from Drosophila suggests that DHR3 inhibits expression of FTZ-F1 in ecdysone regulatory cascades (Kageyama et al., 1997; Lam et al., 1999; White et al., 1997), but no such regulation has been seen in the worm (Kostrouchova et al., 2001). Fly FTZ-F1 has also been shown to form a heterodimeric complex with the FTZ homoedomain transcription factor to regulate embryonic patterning (Guichet et al., 1997; Yu et al., 1997). Similarly, NHR-25 may form a functional complex with two Hox genes, NOB-1 and LIN-39, to influence embryonic and larval cell fates (Chen et al., 2004).
NHR-23 and NHR-25 as well as NHR-41, a DHR78 homolog (Fisk and Thummel, 1998), affect the dauer molt and morphogenesis (Gissendanner et al., 2004). Two other NRs implicated in Drosophila ecdysone cascades have no obvious molting phenotypes. RNAi depletion of NHR-6, the ortholog of DHR38/NGF-IB, instead results in ovulation defects, while knockdown of NHR-85 (E75) causes an egg laying phenotype, suggesting responsibilities in reproductive biology (Gissendanner et al., 2004). Alternately, egg laying defects could result from localized obstruction of cuticle deposition or shedding from the vulva.
Surprisingly, NHR-67 plays an unexpected role in the molt cycle and vulval morphogenesis (Gissendanner et al., 2004). RNAi knockdown results in animals that have difficulty shedding the L3 cuticle and a protruding vulva phenotype, but the cellular basis of these defects remains to be determined. By contrast, the Drosophila homolog TLL has no known role in molting or metamorphosis. Instead it influences anterior/posterior patterning including that of the embryonic CNS (Strecker et al., 1988). The mouse homolog, TLX, functions in the forebrain where it is involved in the generation and differentiation of neurons destined for superficial cortical layers, (Land and Monaghan, 2003; Roy et al., 2002), as well as maintenance of adult neural stem cells (Shi et al., 2004).
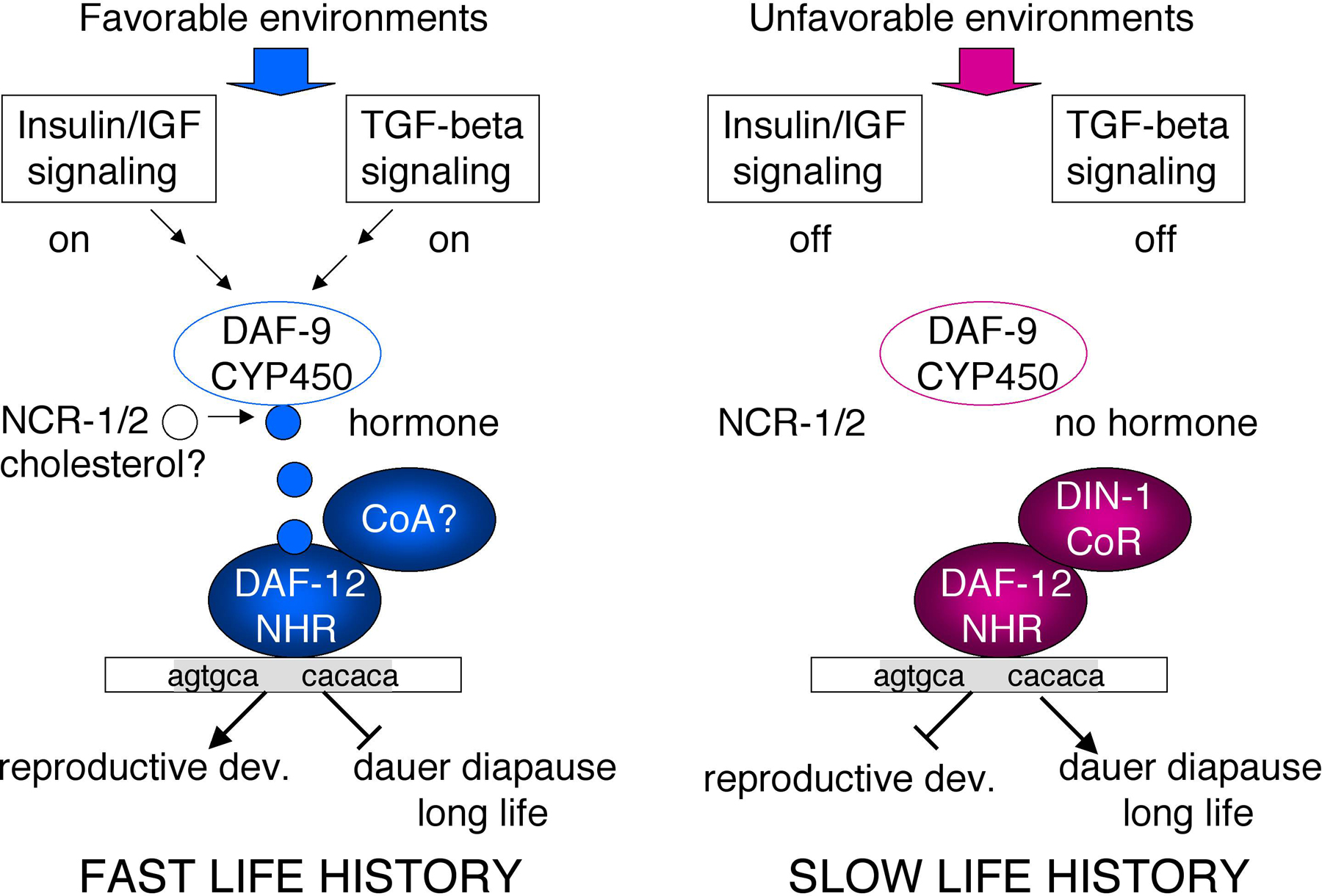
From the perspective of signal transduction, DAF-12 is perhaps the best characterized NR in C. elegans. The outline of an entire hormonal signaling pathway—from signaling inputs, genes involved in hormone transport and metabolism, transcriptional complexes, binding sites, and gene targets—is emerging (Figure 2).
DAF-12 couples environmental cues to life history alternatives, acting at the nexus of pathways governing metabolism, dauer diapause, heterochronic stage selectors, and life span (Antebi et al., 1998; Gerisch et al., 2001; Hsin and Kenyon, 1999; Jia et al., 2002; Larsen et al., 1995; Riddle et al., 1981). For dauer formation, DAF-12 integrates signals from Insulin/IGF-I, TGF-beta and cGMP pathways to mediate either reproductive development or arrest at the dauer diapause. DAF-12 relatives include Vitamin D, Pregnane-X, Liver-X and Androstane receptors (Antebi et al., 2000; Snow and Larsen, 2000), all of which can respond to hormones ultimately derived from cholesterol. Although a DAF-12 hormone has not yet been identified, clear evidence argues for its existence. Notably, DAF-9 encodes a cytochrome P450 related to mixed function oxygenases involved in steroid hormone and xenobiotic metabolism (Gerisch et al., 2001; Jia et al., 2002). daf-9 mutants phenotypically resemble daf-12 LBD mutants: they form dauer larvae constitutively, have heterochronic delays in gonadal outgrowth, and are long lived. Moreover, phenotypes are daf-12(+) dependent. Expressed from a handful of endocrine tissues, DAF-9 works cell non-autonomously to control programs throughout the body (Gerisch and Antebi, 2004; Mak and Ruvkun, 2004).
Preliminary evidence supports the hypothesis that the DAF-12 hormone could be a sterol derivative. Interestingly, cholesterol deprivation produces defects similar to daf-9 and daf-12 LBD mutants (Gerisch et al., 2001, Jia et al., 2002). Moreover, the C. elegans Niemann-Pick C1 homologs, NCR-1 and NCR-2, act at the same point as DAF-9 in the dauer signaling pathways (Li et al., 2004; Sym et al., 2000); such proteins are implicated in intracellular cholesterol trafficking in mammals (Ribeiro et al., 2001). Finally, crude lipid fractions can rescue dauer formation induced by cholesterol starvation (Matyash et al., 2004), as well as the Daf-c phenotypes of daf-9 mutants (Gill et al., 2004; A. Antebi, B. Gerisch, unpublished). Together, these data constitute the first functional evidence for any kind of lipophilic hormone in the worm. Further work should ultimately reveal the molecular identity.
More recently, the coregulator DIN-1 has been shown to bind DAF-12 to specify diapause and long life in the absence of hormone (Ludewig et al., 2004), indicating that the unliganded complex is central to organismal biology. DIN-1 is homologous to human SHARP, a corepressor for nuclear receptors and other transcription factors (Oswald et al., 2002; Shi et al., 2001; Shi et al., 2002). A unifying model is that a DAF-12/coregulator complex works as a hormone regulated switch specifying fast life history in the presence of ligand and slow life history in its absence (Figure 2).
|
Figure 2. Life history regulation by DAF-12 . In favorable environments, Insulin/IGF-I and TGF-beta peptide signal converge on the nuclear receptor branch of the dauer pathways. Niemann-Pick C1 homologs NCR-1/2 deliver cholesterol to DAF-9/cytochrome P450 and perhaps other biosynthetic enzymes, triggering synthesis of a hormone, presumably a sterol. In the presence of hormone, DAF-12/NHR assembles postulated coactivator complexes, and directs expression of genes involved in reproductive development, developmental advance, fat metabolism, and accelerated aging (fast life history traits). In unfavorable environments, hormonal pathways are suppressed. Unliganded DAF-12 together with DIN-1/coregulator specify programs of dauer diapause, delayed development, fat storage, and retarded aging (slow life history traits). Insulin/IGF-I signaling also independently regulates longevity and aspects of diapause.
At least two disparate response elements and several target genes have been identified for DAF-12 (Ao et al., 2004; Shostak et al., 2004). One element (AGTGCA; Shostak et al., 2004) resembles the half sites of VitDR (Freedman et al., 1994) and DHR96 relatives (Fisk and Thummel, 1995). The other (CACACA) is often found juxtaposed to PHA-4/forkhead binding sites in pharyngeal expressed genes, employed in pharyngeal remodeling (Ao et al., 2004). In addition, several key genes in the heterochronic circuit (e.g. lin-28; Antebi et al., 1998; Seggerson et al., 2002) let-7 (Johnson et al., 2003) and dauer pathways (daf-9; Gerisch and Antebi, 2004; Mak and Ruvkun, 2004) are regulated by daf-12, but it is unknown if regulation is direct or indirect. Finally, NHR-23, NHR-25 and NHR-41 also influence dauer morphogenesis and molting (Gissendanner et al., 2004). Understanding their roles in the dauer transcriptional hierarchies will be interesting to pursue.
Xenobiotic defense is key to C. elegans survival given its soil ecology and exposure to plant, fungal and bacterial toxins. Homologous to the xenobiotic receptors CAR and PXR, NHR-8 is proposed to manage xenobiotic stress. Mutants are more sensitive to colchicine and chloroquine (Lindblom et al., 2001). Moreover, NHR-8 is expressed in the worm intestine, the equivalent to the mammalian liver. Xenobiotics often themselves behave as ligands for xenobiotic receptors but whether colchicine, chloroquine or any other compounds stimulate NHR-8 is unknown. Nor is it known whether NHR-8 induces phase 1 and 2 detoxifying enzymes. Several cytochrome P450 enzymes are induced by xenobiotics, but the regulation of this response remains unexplored (Menzel et al., 2001). More recently, NHR-8 has been shown to also influence lipid metabolism since Nile red deposition is altered in RNAi knockdown experiments (Ashrafi et al., 2003).
Vertebrate receptors, such as PPAR, FXR, and LXR, are lipid sensors that regulate fatty acid, bile and cholesterol metabolism. Though orthologs of these receptors are absent in C. elegans, genome-wide screens identified several NRs that increase (NHR-8, NHR-49, C56E10.4, F16B4.9, H12C20.3) or decrease (DAF-12, NHR-25, Y69A2A_7278, C33G8.9, KO8A2.b) lipid deposition when knocked down by RNAi (Ashrafi et al., 2003). Further studies on NHR-49 reveal that it upregulates genes for fatty acid beta oxidation (acs-2, ech-1), desaturation (fat-5, fat-7), and transport (Van Gilst et al., 2004) as well as genes for synthesis of monomethyl branched chain fatty acids (Kniazeva et al., 2004). Mutants accumulate saturated fatty acids and are short lived. Overall, NHR-49 is thought to coordinate fat consumption and the balance of fatty acid saturation. Despite homology to HNF4, NHR-49 may have assumed many of the responsibilities of PPARα, based on its regulatory spectrum (Van Gilst et al., 2004). Other NRs perhaps more similar to HNF4, such as NHR-64 and NHR-69, have no overt phenotype (Gissendanner et al., 2004), but their regulatory spectra have yet to be examined. Interestingly, NHR-49 physically interacts with numerous other NRs by yeast two-hybrid (Li et al., 2004), suggesting it may work as a common heterodimeric partner.
Because the C. elegans nervous system is described down to the synaptic level (White et al., 1986), there is unparalleled opportunity to dissect nematode neurobiology. Several identified NRs affect neural development. UNC-55, an ortholog of the orphan receptor COUP-TF/Seven-up specifies the synaptic wiring of VD motorneurons during the L1stage (Walthall and Plunkett, 1995; Zhou and Walthall, 1998). In mutants, the post-embryonicVD motorneurons differentiate like their embryonic DD counterparts. Expressed in the VD neurons, UNC-55 autonomously prevents expression of the DD fate. Similarly, in the Drosophila eye, Seven-up quells the R7 fate in neighboring photoreceptor cells (Mlodzik et al., 1990). In mice, COUP-TFI affects neural crest ganglionic precursor cells, axon guidance, and early neocortical regionalization (Qiu et al., 1997; Zhou et al., 1999; Zhou et al., 2001). FAX-1 is a homolog of mammalian PNR, a nuclear receptor associated with hyperproliferation of blue cone cells and retinal degeneration (Gerber et al., 2000; Haider et al., 2000). FAX-1 is also related to Tailless/Tlx. Expressed in 20 neurons, FAX-1 alters late aspects of neural fate (Much et al., 2000). In fax-1 mutants, AVK interneurons fail to extend along the ventral cord and into the nerve ring, and fail to express specific neuronal markers. Expression of FAX-1 in AVK suggests a cell autonomous role. However, it also affects the outgrowth of neurons in which it is not expressed (Wightman et al., 1997), suggesting a non-autonomous role in guidepost cells. Recently, FAX-1 has been shown to work in a complementary or combinatorial fashion with the UNC-42/paired-homeodomain protein in specifying aspects of interneuron cell fate (Wightman et al., 2005).
ODR-7 controls the fate of specific olfactory neurons (Sengupta et al., 1996; Sengupta et al., 1994). In mutants, AWA neurons express markers of AWC, suggesting that ODR-7 specifies late aspects of AWA fate, while repressing the AWC fate. Interestingly, ODR-7 lacks an obvious LBD, and the DBD is displaced to the C-terminus. How much of the NR machinery is coopted by this divergent receptor is unclear. odr-7 reveals a surprising level of complexity, with specific residues differently influencing autoregulation, activation, or repression of downstream target genes (Colosimo et al., 2003). Several other divergent NRs are neuronally expressed but their functions are unknown (Miyabayashi et al., 1999).
Undoubtedly the future of NRs lies in the full exploitation of genomic tools available, such as RNAi, two-hybrid, and transcriptional profiling, as well as classical approaches of suppressor and enhancer genetics and transgenesis, to explore regulatory networks. Elucidation of NR signaling may be realized by investigating candidates with similar or contrary phenotypes. In particular, this approach could be used to identify inputs from signal transduction pathways, potential hormone metabolic genes, coactivators and corepressors, and perhaps unknown factors that impinge on receptor activity. By perturbing NR function itself, we may gain further insight into the physiological output by scrutinizing detailed patterns of target gene regulation or metabolic spectra. Furthermore, these analyses may also reveal transcriptional hierarchies, combinatorial control and connectivity with other signaling pathways .
Several specific challenges lie ahead. The dissection of the molting pathways, identification of hormones, the elucidation of lipid metabolic networks, the coordination of neural development, and a detailed exploration of the xenobiotic response come immediately to mind. In addition, little is known about the role of C. elegans NRs in ion balance, stress response, immunity, and numerous other processes where vertebrate receptors have a proven function. With the vast number of C. elegans receptors largely unexplored, many with no obvious phenotype, the challenge will be to discern their physiological responsibility. The other task will be to relate the roles of both ancestral and diverged receptors to functional vertebrate counterparts. Finally, solidifying genetic inferences with biochemistry, e.g. dissecting transcriptional complexes and identifying hormones, will prove crucial to spanning molecular mechanism to physiology.
I am indebted to members of the C. elegans community for communicating results prior to publication, and to members of the Antebi lab for helpful comments on the manuscript.
Aguinaldo, A.M., Turbeville, J.M., Linford, L.S., Rivera, M.C., Garey, J. R., Raff, R.A., and Lake, J.A. (1997). Evidence for a clade of nematodes, arthropods and other moulting animals. Nature 387, 489–493. Abstract Article
Antebi, A., Culotti, J.G., and Hedgecock, E.M. (1998). daf-12 regulates developmental age and the dauer alternative in C. elegans. Development 125, 1191–1205. Abstract
Antebi, A., Yeh, W.H., Tait, D., Hedgecock, E.M., and Riddle, D.L. (2000). daf-12 encodes a nuclear receptor that regulates the dauer diapause and developmental age in C. elegans. Genes Dev. 14, 1512–1527. Abstract
Ao, W., Gaudet, J., Kent, W.J., Muttumu, S., and Mango, S.E. (2004). Environmentally induced foregut remodeling by PHA-4/FoxA and DAF-12/NHR. Science 305, 1743–1746. Abstract Article
Asahina, M., Ishihara, T., Jindra, M., Kohara, Y., Katsura, I., and Hirose, S. (2000). The conserved nuclear receptor Ftz-F1 is required for embryogenesis, moulting and reproduction in C. elegans. Genes Cells 5, 711–723. Abstract Article
Ashburner, M. (1974). Sequential gene activation by ecdysone in polytene chromosomes of Drosophila melanogaster. II. The effects of inhibitors of protein synthesis. Dev. Biol. 39, 141–157. Abstract
Ashrafi, K., Chang, F.Y., Watts, J.L., Fraser, A.G., Kamath, R.S., Ahringer, J., and Ruvkun, G. (2003). Genome-wide RNAi analysis of C. elegans fat regulatory genes. Nature 421, 268–272. Abstract Article
Bourguet, W., Ruff, M., Chambon, P., Gronemeyer, H., and Moras, D. (1995). Crystal structure of the ligand-binding domain of the human nuclear receptor RXR-alpha. Nature 375, 377–382. Abstract Article
Broadus, J., McCabe, J.R., Endrizzi, B., Thummel, C.S., and Woodard, C.T. (1999). The DrosophilaΒ FTZ-F1 orphan nuclear receptor provides competence for stage-specific responses to the steroid hormone ecdysone. Mol. Cell 3, 143–149. Abstract Article
Brooks, D.R., Appleford, P.J., Murray, L., and Isaac, R.E. (2003). An essential role in molting and morphogenesis of C. elegans for ACN-1, a novel member of the angiotensin-converting enzyme family that lacks a metallopeptidase active site. J. Biol. Chem. 278, 52340–52346. Abstract Article
Carmi, I., Kopczynski, J.B., and Meyer, B.J. (1998). The nuclear hormone receptor SEX-1 is an X-chromosome signal that determines nematode sex. Nature 396, 168–173. Abstract Article
Chen, Z., Eastburn, D.J., and Han, M. (2004). The C. elegans nuclear receptor genenhr-25 regulates epidermal cell development. Mol. Cell Biol. 24, 7345–7358. Abstract Article
Colosimo, M.E., Tran, S., and Sengupta, P. (2003). The divergent orphan nuclear receptor ODR-7 regulates olfactory neuron gene expression via multiple mechanisms inC. elegans. Genetics 165, 1779–1791. Abstract
Fayard, E., Auwerx, J., and Schoonjans, K. (2004). LRH-1: an orphan nuclear receptor involved in development, metabolism and steroidogenesis. Trends Cell Biol. 14, 250–260. Abstract Article
Fisk, G.J., and Thummel, C.S. (1995). Isolation, regulation, and DNA-binding properties of three Drosophila nuclear hormone receptor superfamily members. Proc. Natl. Acad. Sci. USA 92, 10604–10608. Abstract
Fisk, G.J., and Thummel, C.S. (1998). The DHR78 nuclear receptor is required for ecdysteroid signaling during the onset of Drosophila metamorphosis. Cell 93, 543–555. Abstract Article
Freedman, L.P., Arce, V., and Perez Fernandez, R. (1994). DNA sequences that act as high affinity targets for the vitamin D3 receptor in the absence of the retinoid X receptor. Mol. Endocrinol. 8, 265–273. Abstract Article
Fu, M., Wang, C., Zhang, X., and Pestell, R.G. (2004). Acetylation of nuclear receptors in cellular growth and apoptosis. Biochem. Pharmacol. 68, 1199–1208. Abstract Article
Gerber, S., Rozet, J.M., Takezawa, S.I., dos Santos, L.C., Lopes, L., Gribouval, O., Penet, C., Perrault, I., Ducroq, D., Souied, E., et al. (2000). The photoreceptor cell-specific nuclear receptor gene (PNR) accounts for retinitis pigmentosa in the Crypto-Jews from Portugal (Marranos), survivors from the Spanish Inquisition. Hum. Genet. 107, 276–284. Abstract Article
Gerisch, B., and Antebi, A. (2004). Hormonal signals produced by DAF-9/cytochrome P450 regulate C. elegans dauer diapause in response to environmental cues. Development 131, 1765–1776. Abstract Article
Gerisch, B., Weitzel, C., Kober-Eisermann, C., Rottiers, V., and Antebi, A. (2001). A hormonal signaling pathway influencing C. elegans metabolism, reproductive development, and life span. Dev. Cell 1, 841–851. Abstract Article
Gill, M.S., Held, J.M., Fisher, A.L., Gibson, B.W., and Lithgow, G.J. (2004). Lipophilic regulator of a developmental switch in C. elegans. Aging Cell 6, 413–421. Abstract Article
Gissendanner, C.R., Crossgrove, K., Kraus, K.A., Maina, C.V., and Sluder, A. E. (2004). Expression and function of conserved nuclear receptor genes in C. elegans. Dev. Biol. 266, 399–416. Abstract Article
Gissendanner, C.R., and Sluder, A.E. (2000). nhr-25, the C. elegans ortholog of ftz-f1, is required for epidermal and somatic gonad development. Dev. Biol. 221, 259–272. Abstract Article
Guichet, A., Copeland, J.W., Erdelyi, M., Hlousek, D., Zavorszky, P., Ho, J., Brown, S., Percival-Smith, A., Krause, H.M., and Ephrussi, A. (1997). The nuclear receptor homologue Ftz-F1 and the homeodomain protein Ftz are mutually dependent cofactors. Nature 385, 548–552. Abstract Article
Haider, N.B., Jacobson, S.G., Cideciyan, A.V., Swiderski, R., Streb, L.M., Searby, C., Beck, G., Hockey, R., Hanna, D.B., Gorman, S., et al. (2000). Mutation of a nuclear receptor gene, NR2E3, causes enhanced S cone syndrome, a disorder of retinal cell fate. Nat. Genet. 24, 127–131. Abstract Article
Hsin, H., and Kenyon, C. (1999). Signals from the reproductive system regulate the lifespan of C. elegans. Nature 399, 362–366. Abstract Article
Hwang, B.J., and Sternberg, P.W. (2004). A cell-specific enhancer that specifies lin-3 expression in the C. elegans anchor cell for vulval development. Development 131, 143–151. Abstract Article
Jetten, A.M., Kurebayashi, S., and Ueda, E. (2001). The ROR nuclear orphan receptor subfamily: critical regulators of multiple biological processes. Prog. Nucleic Acid Res. Mol. Biol. 69, 205–247. Abstract
Jia, K., Albert, P.S., and Riddle, D.L. (2002). DAF-9, a cytochrome P450 regulating C. elegans larval development and adult longevity. Development 129, 221–231. Abstract
Johnson, S.M., Lin, S.Y., and Slack, F.J. (2003). The time of appearance of the C. elegans let-7 microRNA is transcriptionally controlled utilizing a temporal regulatory element in its promoter. Dev. Biol. 259, 364–379. Abstract Article
Kageyama, Y., Masuda, S., Hirose, S., and Ueda, H. (1997). Temporal regulation of the mid-prepupal gene FTZ-F1: DHR3 early late gene product is one of the plural positive regulators. Genes Cells 2, 559–569. Abstract Article
Kniazeva, M., Crawford, Q.T., Seiber, M., Wang, C.Y., and Han, M. (2004). Monomethyl branched-chain fatty acids play an essential role in C. elegans development. PLoS Biol. 2, E257. Abstract Article
Kostrouchova, M., Krause, M., Kostrouch, Z., and Rall, J.E. (1998). CHR3: a C. elegans orphan nuclear hormone receptor required for proper epidermal development and molting. Development 125, 1617–1626. Abstract
Kostrouchova, M., Krause, M., Kostrouch, Z., and Rall, J.E. (2001). Nuclear hormone receptor CHR3 is a critical regulator of all four larval molts of the nematode C. elegans. Proc. Natl. Acad. Sci. USA 98, 7360–7365. Abstract Article
Kuervers, L.M., Jones, C.L., O'Neil, N.J., and Baillie, D.L. (2003). The sterol modifying enzyme LET-767 is essential for growth, reproduction and development in C. elegans. Mol. Genet. Genomics 270, 121–131. Abstract Article
Lam, G., Hall, B.L., Bender, M., and Thummel, C.S. (1999). DHR3 is required for the prepupal-pupal transition and differentiation of adult structures during Drosophila metamorphosis. Dev. Biol. 212, 204–216. Abstract Article
Land, P.W., and Monaghan, A.P. (2003). Expression of the transcription factor, tailless, is required for formation of superficial cortical layers. Cereb. Cortex 13, 921–931. Abstract Article
Larsen, P.L., Albert, P.S., and Riddle, D.L. (1995). Genes that regulate both development and longevity inC. elegans. Genetics 139, 1567–1583. Abstract
Lavorgna, G., Karim, F.D., Thummel, C.S., and Wu, C. (1993). Potential role for a FTZ-F1 steroid receptor superfamily member in the control of Drosophila metamorphosis. Proc. Natl. Acad. Sci. USA 90, 3004–3008. Abstract
Li, S., Armstrong, C.M., Bertin, N., Ge, H., Milstein, S., Boxem, M., Vidalain, P.O., Han, J.D., Chesneau, A., Hao, T., et al. (2004). A map of the interactome network of the metazoan C. elegans. Science 303, 540–543. Abstract Article
Li, J., Brown, G., Ailion, M., Lee, S., and Thomas, J.H. (2004). NCR-1 and NCR-2, the C. elegans homologs of the human Niemann-Pick type C1 disease protein, function upstream of DAF-9 in the dauer formation pathways. Development 131, 5741–5752. Abstract Article
Lindblom, T.H., Pierce, G.J., and Sluder, A.E. (2001). A C. elegans orphan nuclear receptor contributes to xenobiotic resistance. Curr. Biol. 11, 864–868. Abstract Article
Ludewig, A.H., Kober-Eisermann, C., Weitzel, C., Bethke, A., Neubert, K., Gerisch, B., Hutter, H., and Antebi, A. (2004). A novel nuclear receptor/coregulator complex controls C. elegans lipid metabolism, larval development, and aging. Genes Dev. 18, 2120–2133. Abstract Article
Maglich, J.M., Sluder, A., Guan, X., Shi, Y., McKee, D.D., Carrick, K., Kamdar, K., Willson, T.M., and Moore, J.T. (2001). Comparison of complete nuclear receptor sets from the human, C. elegans and Drosophila genomes. Genome Biol. 2, RESEARCH0029. Abstract Article
Mak, H.Y., and Ruvkun, G. (2004). Intercellular signaling of reproductive development by theC. elegans DAF-9 cytochrome P450. Development 131, 1777–1786. Abstract Article
Mangelsdorf, D.J., and Evans, R.M. (1995). The RXR heterodimers and orphan receptors. Cell 83, 841–850. Abstract Article
Mangelsdorf, D.J., Thummel, C., Beato, M., Herrlich, P., Schutz, G., Umesono, K., Blumberg, B., Kastner, P., Mark, M., Chambon, P., and Evans, R.M. (1995). The nuclear receptor superfamily: the second decade. Cell 83, 835–839. Abstract Article
Matyash, V., Entchev, E.V., Mende, F., Wilsch-Brauninger, M., Thiele, C., Schmidt, A.W., Knolker, H.J., Ward, S., and Kurzchalia, T.V. (2004). Sterol-derived hormone(s) controls entry into diapause in C. elegans by consecutive activation of DAF-12 and DAF-16. PLoS Biol. 2, E280. Abstract Article
Menzel, R., Bogaert, T., and Achazi, R. (2001). A systematic gene expression screen of C. elegans cytochrome P450 genes reveals CYP35 as strongly xenobiotic inducible. Arch. Biochem. Biophys. 395, 158–168. Abstract Article
Merris, M., Wadsworth, W.G., Khamrai, U., Bittman, R., Chitwood, D.J., and Lenard, J. (2003). Sterol effects and sites of sterol accumulation in C. elegans: developmental requirement for 4 Α -methyl sterols. J. Lipid Res. 44, 172–181. Abstract Article
Miyabayashi, T., Palfreyman, M.T., Sluder, A.E., Slack, F., and Sengupta, P. (1999). Expression and function of members of a divergent nuclear receptor family in C. elegans. Dev. Biol. 215, 314–331. Abstract Article
Mlodzik, M., Hiromi, Y., Weber, U., Goodman, C.S., and Rubin, G.M. (1990). The Drosophila seven-up gene, a member of the steroid receptor gene superfamily, controls photoreceptor cell fates. Cell 60, 211–224. Abstract Article
Much, J.W., Slade, D.J., Klampert, K., Garriga, G., and Wightman, B. (2000). The fax-1 nuclear hormone receptor regulates axon pathfinding and neurotransmitter expression. Development 127, 703–712. Abstract
Oswald, F., Kostezka, U., Astrahantseff, K., Bourteele, S., Dillinger, K., Zechner, U., Ludwig, L., Wilda, M., Hameister, H., Knochel, W., et al. (2002). SHARP is a novel component of the Notch/RBP-Jkappa signalling pathway. EMBO J. 21, 5417–5426. Abstract Article
Parker, K.L., Rice, D.A., Lala, D.S., Ikeda, Y., Luo, X., Wong, M., Bakke, M., Zhao, L., Frigeri, C., Hanley, N.A., et al. (2002). Steroidogenic factor 1: an essential mediator of endocrine development. Recent Prog. Horm. Res. 57, 19–36. Abstract Article
Prahlad, V., Pilgrim, D., and Goodwin, E.B. (2003). Roles for mating and environment in C. elegans sex determination. Science 302, 1046–1049. Abstract Article
Preitner, N., Damiola, F., Lopez-Molina, L., Zakany, J., Duboule, D., Albrecht, U., and Schibler, U. (2002). The orphan nuclear receptor REV-ERB α controls circadian transcription within the positive limb of the mammalian circadian oscillator. Cell 110, 251–260. Abstract Article
Qiu, Y., Pereira, F.A., DeMayo, F.J., Lydon, J.P., Tsai, S.Y., and Tsai, M. J. (1997). Null mutation of mCOUP-TFI results in defects in morphogenesis of the glossopharyngeal ganglion, axonal projection, and arborization. Genes Dev. 11, 1925–1937. Abstract
Renaud, J.P., Rochel, N., Ruff, M., Vivat, V., Chambon, P., Gronemeyer, H., and Moras, D. (1995). Crystal structure of the RAR- Γ ligand-binding domain bound to all-trans retinoic acid. Nature 378, 681–689. Abstract Article
Ribeiro, I., Marcao, A., Amaral, O., Sa Miranda, M.C., Vanier, M.T., and Millat, G. (2001). Niemann-Pick type C disease: NPC1 mutations associated with severe and mild cellular cholesterol trafficking alterations. Hum. Genet. 109, 24–32. Abstract Article
Riddiford, L.M., Cherbas, P., and Truman, J.W. (2000). Ecdysone receptors and their biological actions. Vitam. Horm. 60, 1–73. Abstract
Riddiford, L.M., Hiruma, K., Zhou, X., and Nelson, C.A. (2003). Insights into the molecular basis of the hormonal control of molting and metamorphosis from Manduca sexta and Drosophila melanogaster. Insect Biochem. Mol. Biol. 33, 1327–1338. Abstract Article
Riddle, D.L., Swanson, M.M., and Albert, P.S. (1981). Interacting genes in nematode dauer larva formation. Nature 290, 668–671. Abstract Article
Robinson-Rechavi, M., Maina, C.V., Gissendanner, C.R., Laudet, V., and Sluder, A. (2005). Explosive lineage-specific expansion of the orphan nuclear receptor HNF4 in nematodes. J. Mol. Evol. in press. Abstract
Roy, K., Thiels, E., and Monaghan, A.P. (2002). Loss of the tailless gene affects forebrain development and emotional behavior. Physiol. Behav. 77, 595–600. Abstract Article
Seggerson, K., Tang, L., and Moss, E.G. (2002). Two genetic circuits repress the Caenorhabditis elegans heterochronic gene lin-28 after translation initiation. Dev. Biol. 243, 215–225. Abstract Article
Sengupta, P., Chou, J.H., and Bargmann, C.I. (1996). odr-10 encodes a seven transmembrane domain olfactory receptor required for responses to the odorant diacetyl. Cell 84, 899–909. Abstract Article
Sengupta, P., Colbert, H.A., and Bargmann, C.I. (1994). The C. elegans geneodr-7 encodes an olfactory-specific member of the nuclear receptor superfamily. Cell 79, 971–980. Abstract
Shi, Y., Chichung Lie, D., Taupin, P., Nakashima, K., Ray, J., Yu, R.T., Gage, F.H., and Evans, R.M. (2004). Expression and function of orphan nuclear receptor TLX in adult neural stem cells. Nature 427, 78–83. Abstract Article
Shi, Y., Downes, M., Xie, W., Kao, H.Y., Ordentlich, P., Tsai, C.C., Hon, M., and Evans, R.M. (2001). Sharp, an inducible cofactor that integrates nuclear receptor repression and activation. Genes Dev. 15, 1140–1151. Abstract Article
Shi, Y., Hon, M., and Evans, R.M. (2002). The peroxisome proliferator-activated receptor delta, an integrator of transcriptional repression and nuclear receptor signaling. Proc. Natl. Acad. Sci. USA 99, 2613–2618. Abstract Article
Shibata, Y., Branicky, R., Landaverde, I.O., and Hekimi, S. (2003). Redox regulation of germline and vulval development in C. elegans. Science 302, 1779–1782. Abstract Article
Shim, Y.H., Chun, J.H., Lee, E.Y., and Paik, Y.K. (2002). Role of cholesterol in germ-line development ofC. elegans. Mol. Reprod. Dev. 61, 358–366. Abstract Article
Shostak, Y., Van Gilst, M.R., Antebi, A., and Yamamoto, K.R. (2004). Identification of C. elegans DAF-12 binding sites, response elements and target genes. Genes Dev. 18, 2529–2544. Abstract Article
Silhankova, M., Jindra, M., and Asahina, M. (2005). Nuclear receptor NHR-25 is required for cell shape dynamics during epidermal differentiation in C. elegans. J. Cell Sci. 118, 223–232. Abstract Article
Sluder, A.E., and Maina, C.V. (2001). Nuclear receptors in nematodes: themes and variations. Trends Genet. 17, 206–213. Abstract Article
Sluder, A.E., Mathews, S.W., Hough, D., Yin, V.P., and Maina, C.V. (1999). The nuclear receptor superfamily has undergone extensive proliferation and diversification in nematodes. Genome Res. 9, 103–120. Abstract
Snow, M.I., and Larsen, P.L. (2000). Structure and expression of daf-12: a nuclear hormone receptor with three isoforms that are involved in development and aging in C. elegans. Biochim. Biophys. Acta 1494, 104–116. Abstract
Stone, B.L., and Thummel, C.S. (1993). The Drosophila 78C early late puff contains E78, an ecdysone-inducible gene that encodes a novel member of the nuclear hormone receptor superfamily. Cell 75, 307–320. Abstract Article
Strecker, T.R., Merriam, J.R., and Lengyel, J.A. (1988). Graded requirement for the zygotic terminal gene, tailless, in the brain and tail region of the Drosophila embryo. Development 102, 721–734. Abstract
Sullivan, A.A., and Thummel, C.S. (2003). Temporal profiles of nuclear receptor gene expression reveal coordinate transcriptional responses during Drosophila development. Mol. Endocrinol. 17, 2125–2137. Abstract Article
Sym, M., Basson, M., and Johnson, C. (2000). A model for Niemann-Pick type C disease in the nematode Caenorhabditis elegans. Curr. Biol. 10, 527–530. Abstract Article
Ueda, H., Sonoda, S., Brown, J.L., Scott, M.P., and Wu, C. (1990). A sequence-specific DNA-binding protein that activates fushi tarazu segmentation gene expression. Genes. Dev. 4, 624–635. Abstract Article
Van Gilst, M.R., Hadjivassiliou, H., Jolly, A., and Yamamoto, K.R. (2004). Nuclear hormone receptor NHR-49 controls fat consumption, storage, and composition in C. elegans. PLoS Biol. 3, e53. Article
Walthall, W.W., and Plunkett, J.A. (1995). Genetic transformation of the synaptic pattern of a motoneuron class in Caenorhabditis elegans. J. Neurosci. 15, 1035–1043. Abstract
White, J.G., Southgate, E., Thomson, J.N., and Brenner, S. (1986). The structure of the nervous system ofC. elegans. Philos. Trans. R. Soc. Lond., B., Biol. Sci. 314, 1–340. Abstract
White, K.P., Hurban, P., Watanabe, T., and Hogness, D.S. (1997). Coordination of Drosophila metamorphosis by two ecdysone-induced nuclear receptors. Science 276, 114–117. Abstract Article
Wightman, B., Baran, R., and Garriga, G. (1997). Genes that guide growth cones along the C. elegans ventral nerve cord. Development 124, 2571–2580. Abstract
Wightman, B., Ebert, B., Jacobs, N., and Clever, S. (2005). The C. elegans nuclear receptor gene fax-1 and homeobox geneunc-42 coordinate interneuron identity by regulating the expression of glutamate receptor subunits and other neuron-specific genes. Dev. Biol. 287, 74–85. Abstract Article
Yochem, J., Tuck, S., Greenwald, I., and Han, M. (1999). A gp330/megalin-related protein is required in the major epidermis of C. elegans for completion of molting. Development 126, 597–606. Abstract
Yu, Y., Li, W., Su, K., Yussa, M., Han, W., Perrimon, N., and Pick, L. (1997). The nuclear hormone receptor Ftz-F1 is a cofactor for the Drosophila homeodomain protein Ftz. Nature 385, 552–555. Abstract Article
Zhou, C., Qiu, Y., Pereira, F.A., Crair, M.C., Tsai, S.Y., and Tsai, M.J. (1999). The nuclear orphan receptor COUP-TFI is required for differentiation of subplate neurons and guidance of thalamocortical axons. Neuron 24, 847–859. Abstract Article
*Edited by Iva Greenwald. Last revised July 5, 2005. Published January 03, 2006. This chapter should be cited as: Antebi, A. Nuclear hormone receptors in C. elegans (January 03, 2006), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.64.1, http://www.wormbook.org.
Copyright: © 2006 Adam Antebi. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
§To whom correspondence should be addressed. E-mail: aantebi@bcm.tmc.edu
 All journal content, except where otherwise noted, is licensed under a Creative Commons Attribution License.
All journal content, except where otherwise noted, is licensed under a Creative Commons Attribution License.