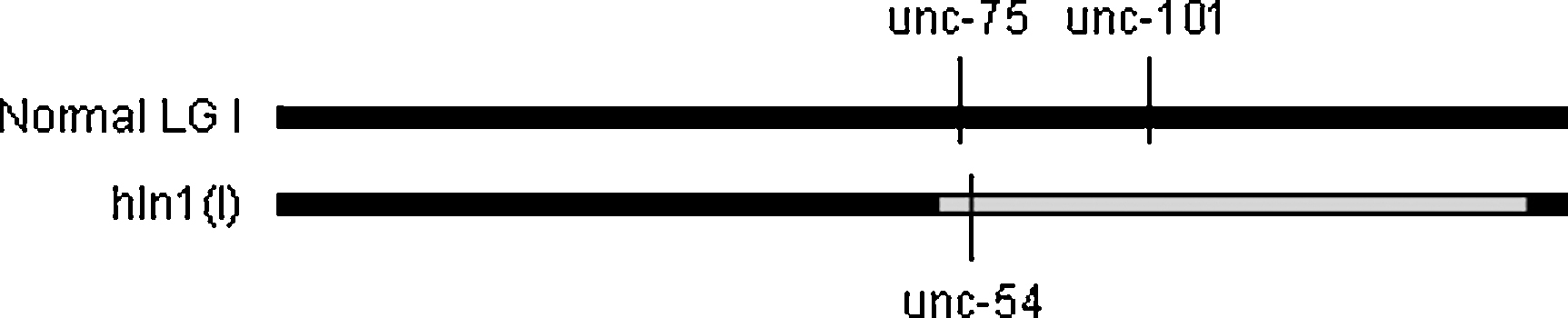Embryo series courtesy of Einhard Schierenberg
Embryo series courtesy of Einhard SchierenbergTable of Contents
Genetic balancers are genetic constructs or chromosomal rearrangements that allow lethal or sterile mutations to be stably maintained in heterozygotes. In this chapter we use the term balancer primarily to refer to chromosomal duplications or rearrangements that suppress crossing over. In addition, we define lethal as any mutation that blocks survival or reproduction. Phenotypes associated with lethal mutations in Caenorhabditis elegans range from egg or larval lethality to adult sterility and maternal effect lethality, and can include conditional effects such as temperature sensitivity. The number of essential genes in C. elegans (those identified by lethal mutations) may range as high as 7000 according to genetic estimates (Clark, D.V., Rogalski, T.M., Donati, L.M., and Baillie, D.L., 1988; Howell, A.M., and Rose, A.M., 1990; Johnsen, R.C., and Baillie, D.L., 1991). Thus, lethal mutations constitute a rich source of information about basic biological processes in this nematode.
This chapter assumes the reader is somewhat familiar with C. elegans genetic nomenclature. A few basic definitions are given here,‡ but for full reference see Horvitz, H.R., Brenner, S., Hodgkin, J., and Herman, R.K. (1979).
Maintenance of mutations is an important aspect of C. elegans genetics. Genetic strains carrying non-conditional recessive lethal mutations, at present the largest class of lethal, cannot be kept as homozygotes, and in heterozygotes the mutations can be lost easily through segregation unless there is a means to identify the heterozygotes that carry them. Maintenance of the heterozygous genotype from one generation to the next requires selection of heterozygous individuals, a task that becomes burdensome if more than a few strains must be maintained. For example, the self progeny of an unmarked recessive lethal heterozygote (let-x/+) are either arrested let-x homozygotes (let-x/let-x), let-x heterozygotes (let-x/+), or wild-type animals (+/+) that cannot be distinguished phenotypically from heterozygotes. Many phenotypically wild-type animals must be selected each generation to ensure the propagation of heterozygotes, and the presence of the lethal mutation must be confirmed through direct observation of let-x homozygotes. If the normal homologue carries a morphological marker, in trans to the lethal mutation, heterozygotes can be reliably identified; however, loss of the lethal can still occur through recombination and subsequent segregation. For example, in a let+/+dpy heterozygote, a crossover between the let and dpy loci produces two recombinant chromosomes, one that carries both mutations and one that is completely wild type. A zygote carrying the wild-type recombinant chromosome and a parental dpy chromosome (+/dpy) will have a wild-type phenotype, and will produce wild-type and Dpy progeny just as the original heterozygote, but the desired lethal will no longer be present in the strain. Thus, the presence of the lethal in each generation still must be confirmed through direct observation or be deduced through the ratio of wild-type to Dpy progeny, which is very tedious (about 2:1 in let+/+dpy heterozygotes and 3:1 in +/dpy heterozygotes).
Clearly, any sort of large-scale isolation and analysis of lethal mutations requires more effective methods to prevent their loss. The investigator must have an easy way to distinguish between progeny heterozygous for a lethal-bearing chromosome and those homozygous for the nonlethal homologue and, further, must be able either to detect directly when recombination has occurred between the lethal mutation and marker mutations or to reduce the incidence of recombination to a negligible level. A balancer provides these functions.
Many different genetic constructs (genotypes) have been used to balance lethal mutations, but they have the same basic characteristics: (1) heterozygotes possess a unique phenotype (wild type or mutant) so that they can be selected reliably; (2) progeny phenotypes allow the investigator to tell when recombination has occurred; (3) close proximity of markers increases the degree of balancing by decreasing the frequency of recombination between lethals and markers; and (4) in the majority of cases, common visible markers linked to the lethals ease all phases of analysis.
Linking lethals to morphological markers is especially helpful for analysis, as it is much easier to score for the absence of a particular morphological phenotype than to score arrested progeny. Furthermore, linked markers facilitate two-factor mapping, and markers common to an entire set of lethals allow rapid inter se complementation tests. For example, recombination frequency can be measured between a lethal mutation and a linked marker mutation by scoring viable marker homozygotes among the progeny segregated from a heterozygote. Complementation between two lethal mutations can be assayed by performing a cross to construct animals heterozygous for both lethals and the same marker mutation, and scoring for marker homozygotes. Presence of these homozygotes indicates complementation, and absence indicates failure to complement.
Three examples of the use of simple genotypes to balance lethal mutations follow. To maintain newly generated deficiencies on linkage group (LG) III that were lethal as homozygotes, Greenwald, I.S., and Horvitz, H.R. (1980) kept them as heterozygotes over an unc-93 dpy-17 chromosome. These animals were Unc-93 in phenotype because the deficiencies deleted unc-93, and they segregated Unc-93, Unc-93 Dpy-17, and lethal (deficiency homozygote) progeny. The deficiencies could be maintained by picking Unc animals, as the recombination frequency between dpy-17 and the deficiency endpoints was low. In a second example, Moerman, D.G., and Baillie, D.L. (1979) isolated lethals linked to a conditionally dominant unc-22 mutation, which causes dominant twitching in the presence of 1% nicotine. Conditional dominance results in a unique heterozygous phenotype. In screens for new mutations, worms heterozygous for unc-22 were screened for the absence or near absence of fertile adult Unc-22 progeny. Once obtained, these lethals could be maintained by picking animals that moved normally but were induced to twitch in 1% nicotine. Crossing over between a lethal and unc-22 was easily detected by scoring the rare Unc-22 progeny in the absence of nicotine. The closer the lethal was to unc-22, the less often recombinational loss occurred. In a third example, Rose, A.M., and Baillie, D.L. (1980) mutagenized a phenotypically wild-type strain of genotype +unc-15+/dpy-14+unc-13 and screened for lethal mutations linked to the dpy-14 unc-13 Ichromosome. The new lethals were maintained simply by picking wild-type animals (heterozygotes) and checking to see that they gave wild-type and Unc-l5 progeny but no Dpy-14 Unc-13 progeny. Lethals were balanced because of the short genetic distances. In all cases, recombination was easily detected by the presence of Dpy, Unc, or DpyUnc recombinant animals among the self progeny of a heterozygote. The types and numbers of recombinant progeny gave distance and gene order relative to the markers.
A drawback of these methods is that they are not easily adapted for the isolation of large numbers of lethal mutations over large genetic regions. They may require many different sets of appropriate marker mutations, which either may be too difficult to score or may not exist. A more sophisticated approach makes use of heterozygous chromosomal rearrangements adapted for use as balancers. There are two types of balancing rearrangements: (1) those that reduce or eliminate recombination between a lethal-bearing chromosome and a homologue carrying a wild-type allele of the locus, and (2) those that provide an extrachromosomal or integrated wild-type allele that complements a homozygous lethal mutation. As balancers, these rearrangements have several advantages over simple marked chromosomes. They can balance large genomic regions, and the choice of markers to which lethals can be linked is much broader, requiring only that the markers lie in the balanced region. By virtue of their effects on recombination, they can be heritably stable vehicles for the maintenance of large numbers of lethal mutations. Finally, genetic variants of the rearrangements themselves can be generated to aid in strain construction, mutant screens, and analysis of balancer structure.
Rearrangements as balancers are the focus of this chapter. We describe the features and behavior of each class of balancer currently used in C. elegans, using a well-characterized member of the class; present a discussion of the practical aspects of day-to-day balancer use; give an overview of the state of balancing for each chromosome; and lay out in guidebook form information for understanding and maintaining the most commonly used C. elegans balancers and their genetic variants. A great deal of detailed analysis beyond the scope of this chapter has been accumulated on balancers (see citations for individual rearrangements), and we encourage the reader to seek out the published works for a more complete understanding of the behavior of particular balancers. Information contained in the cited work is especially helpful when designing mutant screens and carrying out complementation and mapping protocols.
In C. elegans, Herman, R.K., Albertson, D.G., and Brenner, S. (1976) first characterized duplications with the intention of using them to recover recessive lethal mutations on the X chromosome. Subsequent work by many different groups has resulted in the isolation and characterization of an array of rearrangement types that together balance approximately 70% of the C. elegans genome. These rearrangements, including translocations, duplications, and inversions, constitute a powerful set of tools for investigators. In addition, other dominant crossover suppressors and transgenes have been adapted for use as balancers. The physical structures of several balancers have been deduced through genetic analysis. Recent work using fluorescence in situ hybridization (FISH) analysis (Albertson, D.G., Fishpool, R.M., and Birchall, P.S., 1995) shows the potential for determining balancer structure through molecular means. The following discussion gives an overview of the different classes of balancers.
Translocations are chromosomal rearrangements in which parts of nonhomologous chromosomes are exchanged (Figure 1). Translocations in C. elegans have been recovered in specific screens for X-chromosome nondisjunction (Herman, R.K., Kari, C.K., and Hartman, P.S., 1982), suppression of recombination between widely spaced linked markers (Herman, R.K., 1978; Fodor, A., and Deak, P., 1985; Edgley, M.L., and Riddle, D.L., 2001), linked lethals (Rosenbluth, R.E., Cuddeford, C., and Baillie, D.L., 1985; McKim, K.S., Peters, K., and Rose, A.M., 1993), and pseudolinkage of normally unlinked marker mutations (McKim, K.S., Howell, A.M., and Rose, A.M., 1988; McKim, K.S., Peters, K., and Rose, A.M., 1993). A number of existing translocations have been characterized genetically, and several have proven to be reliable balancers for lethal mutant screens (eT1: Rosenbluth, R.E., and Baillie, D.L., 1981; Rosenbluth, R.E., Cuddeford, C., and Baillie, D.L., 1983; nTl: Clark, D.V., Rogalski, T.M., Donati, L.M., and Baillie, D.L., 1988; hTl: Howell, A.M., and Rose, A.M., 1990). They balance large genomic regions, and they are easily manipulated in genetic crosses.
|
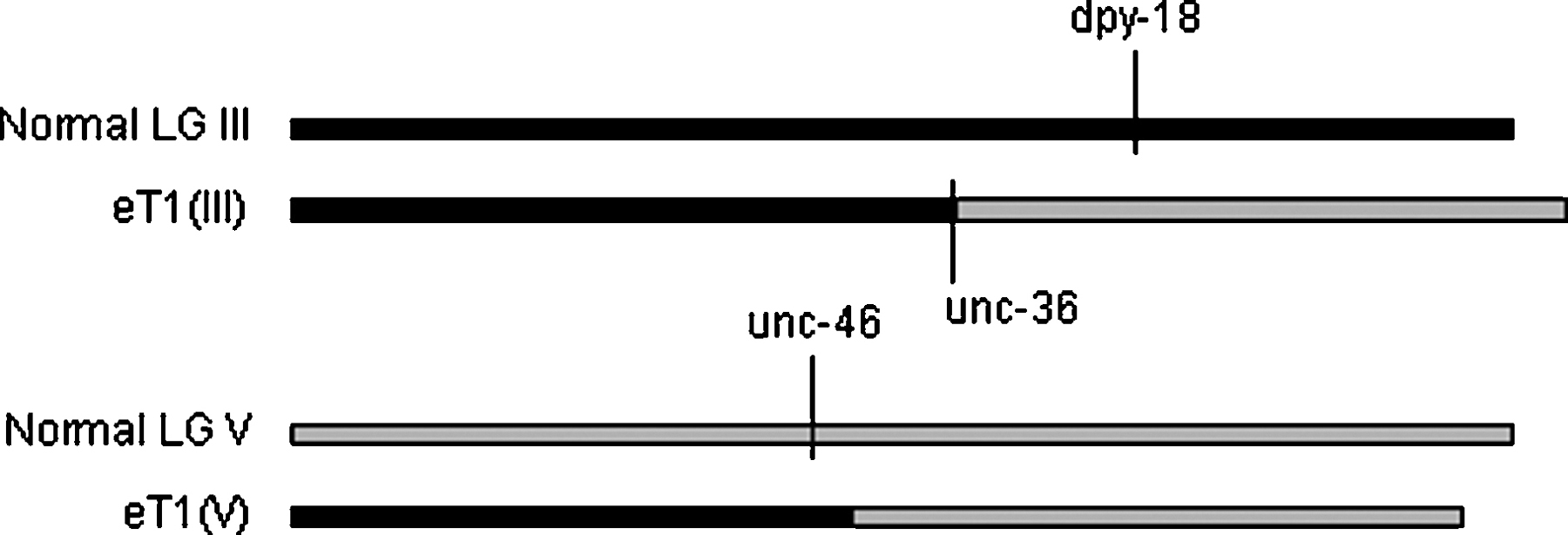
Figure 1. Diagram of reciprocal translocation eT1, showing mutations present in the reference heterozygous strain. The dpy-18 and unc-46 mutations are carried on the normal chromosomes, and the unc-36 mutation is caused by the breakpoint of the translocation on LG III. Recombination is suppressed in the region to the right of unc-36 on LG III, balancing dpy-18, and in the region to the left of unc-42 on LG V (not shown, see Figure 7), balancing unc-46. eT1(III) recombines with the normal LG III in the region to the left of unc-36. eT1(V) recombines with the normal LG V in the region to the right of unc-23 (not shown, see Figure 7).
The first C. elegans translocation for which reciprocal exchange of chromosomal segments was demonstrated is eT1(III;V) (Rosenbluth, R.E., and Baillie, D.L., 1981), which is viable as a homozygote and was originally called unc-72 (Brenner, S., 1974). Marker mutations crossed onto or induced on eT1 were employed to show linkage between appropriate normally unlinked markers on both half-translocations. Several other translocations have since been shown to be reciprocal using similar protocols, for example, mnT2 and mnT10 (Herman, R.K., Kari, C.K., and Hartman, P.S., 1982); szT1 and hT1 (McKim, K.S., Howell, A.M., and Rose, A.M., 1988); and hT2 (McKim, K.S., Peters, K., and Rose, A.M., 1993). Segregation data for nT1 (Ferguson, E.L., and Horvitz, H.R., 1985; Clark, D.V., Rogalski, T.M., Donati, L.M., and Baillie, D.L., 1988), hT3 (McKim, K.S., Peters, K., and Rose, A.M., 1993) and mT1 (Edgley, M.L., and Riddle, D.L., 2001) are consistent with their being reciprocal translocations, but rigorous proof is lacking.
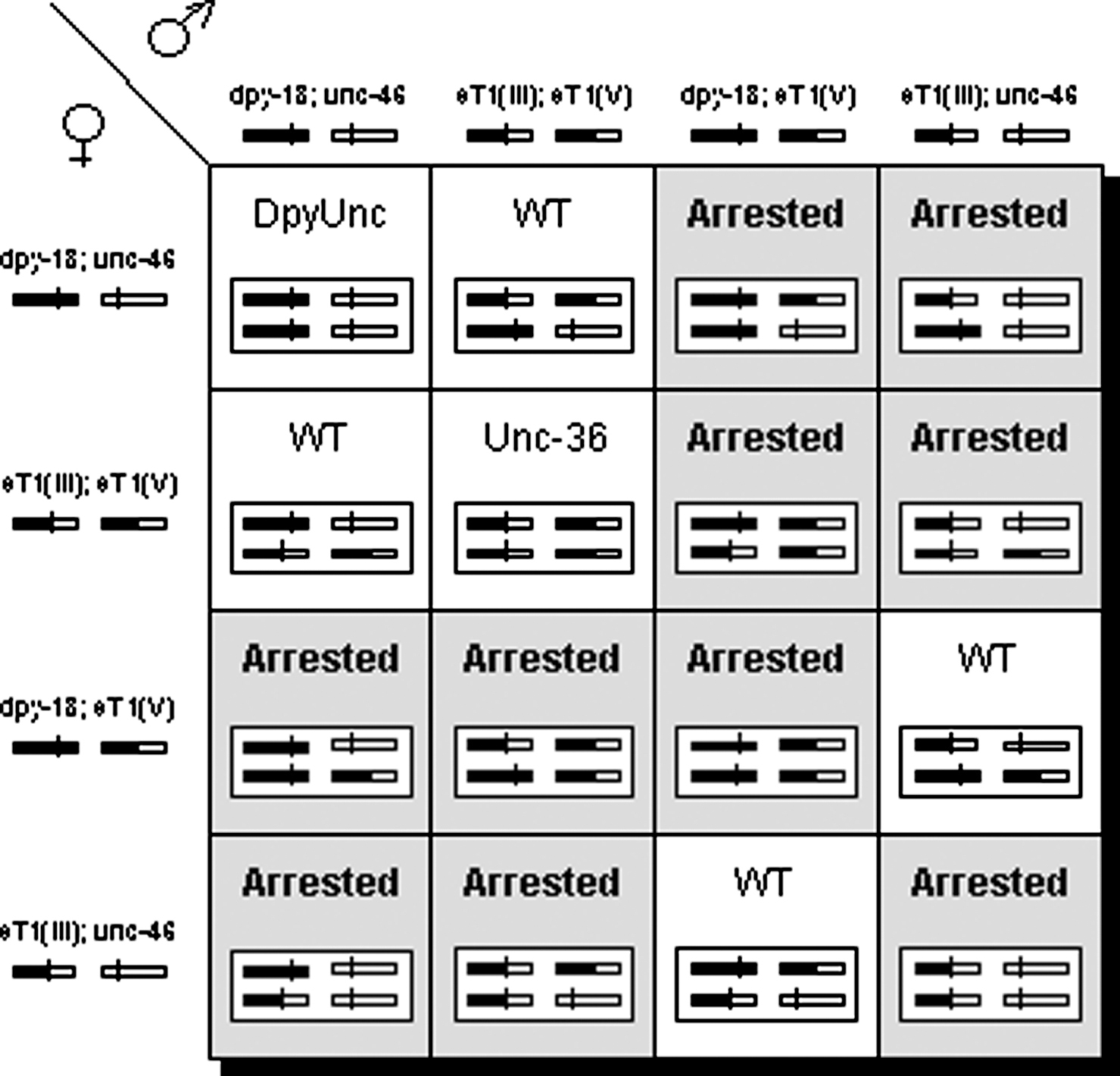
In the case of eT1, the left portion of LG V is translocated to the left portion of LG III, and the right portion of III is translocated to the right portion of V (Figure 1). In heterozygotes, the half-translocation carrying the left portion of III [referred to as eT1(III)] recombines with the normal III in the region to the left of unc-36, and the half-translocation carrying the right portion of V [eT1(V)] recombines with the normal V in the region to the right of unc-23 (refer to Figure 7b and Figure 7c for position of markers). The regions that recombine segregate from each other during meiosis (Rosenbluth, R.E., and Baillie, D.L., 1981; McKim, K.S., Howell, A.M., and Rose, A.M., 1988; McKim, K.S., Peters, K., and Rose, A.M., 1993). Thus, eT1(III) segregates from the normal III, and eT1(V) segregates from the normal LG V. Recombination is suppressed along the length of the translocated portion of each chromosome, from unc-36 to the right end of LG III, and from between unc-23 and unc-42 to the left end of LG. V. The boundaries of crossover suppression thus correspond to the translocation breakpoints. Recombination is not suppressed in the regions that segregate from their normal homologues, from unc-36 to the left end of LG III, and from between unc-23 and unc-42 to the right end of LG V; nor is it suppressed in eT1 homozygotes. These observations have led to the proposal that these regions do not recombine in heterozygotes due to their inability to pair with normal homologues (Rosenbluth, R.E., and Baillie, D.L., 1981). To maintain the translocation as a heterozygote, the normal homologues are marked with morphological mutations in the recombination-suppressed region [in the case of eT1, the most frequently used strain carries dpy-18(III) and unc-46(V)]. In this way, heterozygotes (which have a wild-type phenotype) can be distinguished from normal homologue homozygotes. Self progeny of eT1 heterozygotes are wild-type heterozygotes; eT1 homozygotes (which have an Unc-36 phenotype because the translocation breakpoint lies in the unc-36 gene on LG III); Dpy-18 Unc-46 homozygotes; and a large percentage of aneuploid progeny (10/16) that arrest development (Figure 2) as embryos or early larvae (Adames, K.A., Gawne, J., Wicky, C., Muller, F., and Rose, A.M., 1998; Turner and Baillie, unpublished results). Aneuploid progeny are those with an abnormal complement of chromosomes. Figure 2 illustrates how they arise. The unc-46 and dpy-18 mutations appear to be linked (they are pseudolinked) because all singly mutant homozygotes are aneuploid, and this arrangement cannot break down in the absence of recombination. If a strain carries a recessive lethal mutation in the recombination-suppressed region of either normal homologue, all of the homozygous DpyUnc progeny will die. To characterize these arrested animals, they must not be confused with aneuploid progeny segregated by translocation heterozygotes (See 3.6.6, Linked markers, for more detailed discussion).
|
Figure 2. Punnett square showing normal progeny genotypes and phenotypes that result from selfing a dpy-18/eT1 III; unc-46/eT1 V heterozygote. Phenotypes are indicated for each progeny class. Genotypes for gametes and zygotes are given in gene and rearrangement names according to standard nomenclature, and in drawings representing normal and translocation chromosomes with genetic markers in place. The normal LG III is shown as a black bar, with a vertical line indicating the position of the dpy-18 mutation. The normal LG V is shown as a shaded bar, with a vertical line indicating the position of the unc-46 mutation. The half-translocation eT1 (III) is shown as a half-black, half-shaded bar with a vertical line indicating the position of the unc-36 mutation caused by the translocation breakpoint on LG III. The half-translocation eT1 (V) is shown as a half-black, half-shaded bar with no vertical line. Boxes of the square representing viable progeny are unshaded. Boxes representing aneuploid progeny, all of which arrest during development, are shaded. All wild-type progeny are heterozygous for the translocation chromosomes and the normal chromosomes. unc-36 progeny are eT1 homozygotes, and Dpy Unc progeny are dpy-18; unc-46 homozygotes. Aneuploid progeny account for 10/16ths of the total.
Duplications are chromosomal segments that are present in the nucleus in addition to a full complement of normal chromosomes (Figure 3). Duplications have been generated by mutagenesis with ionizing radiation or UV light, and recovered as genetic elements that rescue the phenotypic expression of particular mutations (Herman, R.K., Albertson, D.G., and Brenner, S., 1976 Rosenbluth, R.E., and Baillie, D.L., 1981; Herman, R.K., Madl, J.E., and Kari, C.K., 1979; Herman, R.K., Kari, C.K., and Hartman, P.S., 1982; Hodgkin, J., 1980; Greenwald, I.S., Sternberg, P.W., and Horvitz, H.R., 1983; Herman, R.K., 1984; Herman, R.K., 1987; Meneely, P.M. and Wood, W.B., 1984; Rose, A.M., Baillie, D.L., and Curran, J., 1984; Rosenbluth, R.E., Cuddeford, C., and Baillie, D.L., 1985; Rosenbluth, R.E., Rogalski, T.M., Johnsen, R.C., Addison, L.M., and Baillie, D.L., 1988; Austin, J., and Kimble, J., 1989; DeLong, L., Casson, L.P., and Meyer, B.J., 1987; Meneely, P.M., and Nordstrom, K.D., 1988; Rogalski, T.M., and Riddle, D.L., 1988; Herman, R.K., and Kari, C.K., 1989; Howell, A.M., and Rose, A.M., 1990; Hunter, C.P., and Wood, W.B., 1990; McKim, K.S., and Rose, A.M., 1990; Yuan, J.Y., and Horvitz, H.R., 1990; Stewart, H.I., Rosenbluth, R.E., and Baillie, D.L., 1991; Marra, M.A., and Baillie, D.L., 1994). They have also been recovered as exceptional segregants from translocation strains (mnDp11: Herman, R.K., Kari, C.K., and Hartman, P.S., 1982; szDp1: McKim, K.S., Howell, A.M., and Rose, A.M., 1988; hDp133, hDp134, hDp135: McKim, K.S., Peters, K., and Rose, A.M., 1993), and as products of a rare recombination event in an inversion strain (Zetka, M.C., and Rose, A.M., 1992). Most duplications in C. elegans are "free" elements that segregate in a non-Mendelian fashion. They have been described as extrachromosomal fragments of genetic material, and they can be lost both mitotically and meiotically during gametogenesis (Herman, R.K., Albertson, D.G., and Brenner, S., 1976). Mitotic chromosomes exhibit multiple spindle attachment points (Albertson, D.G., and Thomson, J.N., 1982), whereas meiotic chromosomes exhibit a localized spindle attachment point (Albertson, D.G., and Thomson, J.N., 1993). Hence, mitotic segregation of chromosome fragments is more easily understood than is meiotic segregation. The mechanism of free duplication transmission during meiosis is unknown. Each duplication has a characteristic frequency of transmission to gametes. Generally, the larger the duplication, the more stable it is in the germ line (McKim, K.S., and Rose, A.M., 1990). Mitotic loss of certain free duplications from somatic cells during development has made possible mosaic analysis of the gene function (Herman, R.K., 1995). A small percentage of duplications exist as integrants into otherwise normal chromosomes, in which case segregation becomes Mendelian. Many duplications have been used as balancers (e.g., mnDpl: Meneely, P.M., and Herman, R.K., 1979; sDp2: Howell, A.M., Gilmour, S.G., Mancebo, R.A., and Rose, A.M., 1987; qDp3: Bucher, E.A., and Greenwald, I., 1991; sDp3: Stewart and Baillie, unpublished results). There are two basic classes of duplication, those that do not recombine with the normal homologues and those that do. Discussion of the significance of this observation can be found in Herman, R.K., Madl, J.E., and Kari, C.K. (1979) and Rose, A.M., Baillie, D.L., and Curran, J. (1984). Members of the nonrecombining class can be used as effective balancers (Rose, A.M., Baillie, D.L., and Curran, J., 1984; Stewart and Baillie, unpublished results). For example, sDp2 has been used extensively in screens for lethal mutations, and it has been shown to permit identification of the same range of essential genes as does a translocation (hT1) that balances the same region (McKim, K.S., Howell, A.M., and Rose, A.M., 1988). Rarely, free duplications have been observed to shorten spontaneously, which reduces the extent of the balanced region (McKim, K.S., and Rose, A.M., 1990).
Free duplications generally exist as a single copy, but strains carrying two copies have been documented in some cases (Herman, R.K., Madl, J.E., and Kari, C.K., 1979; Rogalski, T.M., and Riddle, D.L., 1988). The reference strain carrying sDp2 is homozygous for mutant alleles of dpy-5 and unc-13 and, in addition, has a single copy of sDp2 (Figure 3). The strain has an Unc-13 phenotype, as the duplication carries a wild-type allele of dpy-5 but not of unc-13. It segregates Unc-13 progeny (carrying the duplication) and Dpy-5 Unc-13 homozygotes. The possibility of losing either marker is eliminated by the homozygous state of the marked chromosomes and lack of recombination with the duplication. If a strain is also homozygous for a lethal mutation on the same chromosome in the duplicated region, then the Dpy Unc progeny class will die and the only viable progeny will be those carrying the duplication. The lethal mutation is thus effectively balanced.
|
Figure 3. Diagram of genotype of the reference sDp2 strain, showing progeny genotypes and phenotypes that result from selfing. The strain is homozygous for dpy-5 and unc-13 mutations, and carries one copy of sDp2. Animals of this genotype are Unc-13 in phenotype. Progeny not carrying the duplication are Dpy-5 Unc-13.
A transgene is a segment of cloned DNA containing the complete coding element and control elements of a gene, present in the nucleus in addition to a complete complement of normal chromosomes. Direct transformation of mutant strains using cloned DNA is performed in C. elegans by germline microinjection (Fire, A., 1986; Mello, C., and Fire, A., 1995). The microinjected molecules (containing one or more transgenes) assemble into tandem arrays that can be maintained extrachromosomally. Expression of wild-type transgenes in the array can complement, or "rescue," a mutant phenotype. An important advance in these procedures involved coinjecting the DNA to be tested for rescue along with clones of a selectable marker mutation, such as the dominant rol-6 mutation (Mello, C.C., Kramer, J.M., Stinchcomb, D., and Ambros, V., 1991), which causes animals carrying it to roll along their long axes. Transgenic progeny displaying the marker phenotype were scored for rescue of the strain's mutant phenotype.
Germline microinjection becomes laborious if one wishes to assay the rescuing ability of a group of cosmids in a set of mutants. A more general approach takes advantage of the ability to manipulate transgenes genetically, as if they were free duplications. McKay, McDowall, and Rose (unpublished results) have used transgenes as genetic elements in large-scale experiments to rescue lethal mutations in the dpy-5 and dpy-14 regions of chromosome I. Copies of a marker plasmid containing rol-6(su1006) were coinjected with sets of two or three overlapping cosmids from the genomic region of interest into wild-type animals or animals homozygous for a morphological mutation. Transformants were selected on the basis of their Rol-6 phenotype, and individual lines were established that exhibited a high frequency of transmission of the transgene. Standard duplication complementation tests were then used to assay for rescue of a mutation. Rescue was confirmed by establishment of a nonmutant line from the complementation test that exhibited the Rol-6 phenotype, with the transgene as an effective balancer, and that segregated nonrolling progeny (not carrying the transgenic array) that were not rescued. Since the sequence of the whole genome has been deduced (C. elegans Sequencing Consortium, 1998), this method can be used with virtually any gene.
Complementation tests with extrachromosomal arrays containing more than one cosmid increase the efficiency of the analysis, as every mutation does not have to be tested separately against every cosmid. If the transgenic arrays are properly designed, a small number of crosses can establish exactly which individual cosmid is responsible for the rescue. Arrays containing single rescuing cosmids can then be generated to confirm the result.
Transgenic arrays have also been used as balancers in screens for new mutations. Labouesse and Horvitz (pers. comm.) used a transgenic strain carrying a cosmid that rescues lin-26 in a noncomplementation screen for new mutations mapping to the region balanced by the cosmid. A wild-type clone of dpy-10 was coinjected with the cosmid into a dpy-10 mutant strain as a marker to identify transformants. The resulting non-Dpy transgenic animals were mutagenized, and their F2 progeny screened for the presence of new lin-26 mutations or other mutations balanced by the cosmid. These were identified in the Dpy segregants that had spontaneously lost the transgenes. One new mutation, in the gene let-253, was recovered among the progeny of 2727 F1's. This technique allows balancers to be used in tightly focused screens for mutations in very small genomic regions. Plasterk, R.H.A. (1995) also used such arrays to screen for lethal transposon-induced deficiencies.
Green fluorescent protein (GFP) transgenes (Chalfie M., Tu, Y., Euskirchen, G., Ward W.W., and Prasher, D.C., 1994) also have utility as balancers and chromosome markers. When animals are transformed by microinjection of the GFP coding region under control of a C. elegans promoter, GFP is expressed in the corresponding tissue. For example, a myo-2::GFP construct results in GFP expression in the pharynx. If such a transgene is integrated into the genome, GFP expression becomes a marker for the chromosome into which the element is integrated. The expression of myo-2::GFP constructs typically is semi-dominant, allowing one integrated copy to be distinguished from two by the relative brightness of the GFP signal (Edgley, M.L., and Riddle, D.L., 2001; Edgley, unpublished results).
Several researchers have performed integration experiments with a myo-2::GFP construct, resulting in GFP-tagged variants of several existing balancers and many integrations into non-balancer chromosomes. The former group is very useful, often allowing unequivocal identification of lethal homozygotes by virtue of the absence of GFP expression (see the handbook section for GFP variants of mIn1, nT1, and hT2). Simple chromosomal insertions are also valuable, as they are semi-dominant markers for particular chromosome regions, and they usually exhibit local recombination suppression over a few map units. A number of these elements have been roughly mapped genetically and are in use as balancers for regions where no other balancer exists (Edgley, unpublished results). Large-scale integration of promoter::GFP elements and subsequent SNP mapping could yield local balancer elements for several regions that are not currently covered by balancers.
Inversions are chromosome segments that break from the intact chromosome and rejoin it in the original location but in opposite orientation. There are two proven examples of inversion in C. elegans, hIn1 I and mIn1 II. In hIn1 (Figure 4) a portion of the right half of LG I, from about unc-75 to about unc-54, is inverted (Zetka, M.C., and Rose, A.M., 1992). In mIn1, a large section in the middle of the chromosome is inverted, from about lin-31 to rol-1. For both elements, mapping of mutations in inversion homozygotes revealed the inverted gene order relative to wild type. Both were recovered in screens for rearrangements that suppress crossing over specifically in previously unbalanced regions. Recombination between the breakpoints is almost completely suppressed in heterozygotes, presumably because of absence of DNA alignment or interruption of sites required for pairing. Both inversions are viable as homozygotes, with normal brood sizes, and in the homozygous state exhibit normal levels of recombination in the inverted interval. hIn1 has been used in screens for lethal mutations on LG I to the right of unc-75 (Ho and Rose, unpublished results); mIn1 has been used in screens to isolate lethals and deletions across its extents, especially to the left of dpy-10.
|
Figure 4. Diagram of genotype of reference hIn1 inversion strain. The unc-75 and unc-101 mutations are carried on the normal LG I. The unc-54 mutation was induced secondarily on the inversion chromosome. Recombination is suppressed along the length of the inverted segment in heterozygotes (boundaries of suppression correspond to breakpoints). Terminal sequences are most likely not included in the inverted interval, as a deficiency of the ribosomal gene cluster (eDf24) to the right of unc-54 on normal LG I is apparently not balanced.
The original hIn1 was generated on a wild-type chromosome, making it impossible to distinguish between inversion heterozygotes and homozygotes. To circumvent this difficulty and to make the balancer more useful for mutant screens, new unc-75 (two lethal alleles), unc-54, and unc-101 mutations were induced on hIn1 (Lee, J., Jongeward, G.D., and Sternberg, P.W., 1994; Zetka, M.C., and Rose, A.M., 1992). The reference strain is a phenotypically wild-type heterozygote in which an hIn1[unc-54] chromosome balances unc-75 unc-101. Selfing the strain produces wild-type heterozygotes, balancer homozygotes (Unc-54 phenotype), and Unc-75 Unc-101 homozygotes. hIn1 may not include the physical end of the chromosome, as eDf24 (a deletion of part of the ribosomal gene cluster that lies to the right of unc-54) apparently is not balanced by the inversion (M. Zetka, pers. comm.).
The mIn1 inversion was generated on a dpy-10 chromosome. Several marker variants, including unmarked, doubly-marked, and marked with rol-1 were used to establish gene order. In addition, a variant carrying an integrated pharyngeal GFP in addition to dpy-10 was generated by gamma-ray mutagenesis.
Several chromosomal rearrangements exist in C. elegans that dominantly suppress crossing over but that remain uncharacterized with regard to their structures. Two examples are mnC1 (Herman, R.K., 1978), which balances the right half of LG II from dpy-10 to around unc-52, and sC1 (Stewart and Baillie, unpublished results), which balances an approximately 15-map-unit segment of the left half of LG III, from around unc-45 to the region left of dpy-17. Crossover suppression in these two cases is presumably restricted to a single chromosome, in contrast to translocations. The patterns of crossover suppression, meiotic properties, brood size, and progeny phenotypes are consistent with their being inversions, but proof is lacking. Both have been used successfully in screens for lethal mutations in the regions they balance. mnC1, marked with dpy-10 and unc-52 mutations, has been especially useful in isolating and maintaining a high-resolution set of deficiencies (Sigurdson, D.C., Spanier, G.J., and Herman, R.K., 1984). It is phenotypically wild type as a heterozygote, gives broods of relatively normal size, and is viable as a DpyUnc homozygote but with very small broods. sC1 was generated on a wild-type chromosome, is viable as a homozygote, and morphologically marked variants have been generated to facilitate its use (Stewart and Baillie, unpublished results).
Deficiencies are segments of chromosomes entirely missing from the genome. Deficiencies are detected after mutagenesis by various methods (e.g., Riddle, D.L., and Brenner, S., 1978; Sigurdson, D.C., Spanier, G.J., and Herman, R.K., 1984; Moerman, D.G., and Baillie, D.L, 1981; Greenwald, I.S., and Horvitz, H.R., 1980; Rosenbluth, R.E., Cuddeford, C., and Baillie, D.L., 1985; Yandell, M.D., Edgar, L.S., and Wood, W.B., 1994). They are useful as genetic mapping tools, but some deficiencies dominantly reduce recombination frequency in particular genetic regions. Several of the latter class were described by Rosenbluth, R.E., Johnsen, R.C., and Baillie, D.L. (1990). A chromosome carrying a small deficiency in the gene cluster of LG I, hDf8, was also associated with reduced recombination frequency, from around dpy-14 to around the left end of the chromosome (McKim, K.S., Starr, T., and Rose, A.M., 1992).
Balancers are used for a variety of tasks, including balancing existing mutations, facilitating strain construction, and screening for new mutations. At a minimum, a balancer should have the properties already described: heterozygotes must have a phenotype distinguishable from that of each homozygote; recombination should be virtually eliminated; progeny phenotypes must allow ready detection of rare recombination; and the construct should be stable (balanced mutations should not spontaneously become unbalanced). It is also helpful if the balancer can be manipulated easily in genetic crosses, particularly if it can be passed through male sperm, and if morphologically marked or lethal variants exist.
Balancing existing mutations is probably the least demanding of these tasks, requiring only that a suitable balancer exists and that the mutation/balancer heterozygote is viable. A minor degree of instability in the heterozygote is acceptable, as long as the investigator checks the strain routinely. Strain construction can be aided by particular qualities of a few balancers, and often requires balancers marked with particular mutations (see Section 3.3).
By far the most stringent test of a balancer is to use it to screen for lethal mutations. The major additional requirement imposed on the balancer is that the apparent spontaneous mutation frequency should be low (no higher than in the reference wild-type strain, N2). Studies using two different reciprocal translocations (eT1 and szT1) illustrate this point.
1. eT1(III;V) This translocation is an extremely stable balancer that has been used extensively for lethal screens. The size of the balanced region is about 40 map units (mu). Rosenbluth, R.E., Cuddeford, C., and Baillie, D.L. (1983) conducted a dose-response study for balanced lethal mutations using ethyl methanesulfonate (EMS) and gamma irradiation, and determined the spontaneous mutation frequency for lethal mutations. A strain of genotype dpy-18/eT1 III; unc-46/eT1 V was used to screen for new lethal mutations linked to either dpy-18 or unc-46. Lethals were recovered at reasonable frequency with both mutagens (6.6% at 12 mM EMS, and 4.5% at 1500 R gamma). The frequency of accumulation of spontaneous mutations was very low. Two control experiments, involving 3198 F1 heterozygous progeny of two P0 animals, resulted in a total of two balanced lethal mutations for a frequency of 0.06%. In addition, eT1 seems to be completely stable. It has been used to screen many thousands of mutagenized chromosomes, is currently used to balance hundreds of lethal mutations, and it has never been observed to break down.
2. szT1(I;X) This translocation is very stable, and it has been useful in strain constructions and for balancing existing mutations. It has been used to a limited extent to recover new lethal mutations. Experiments to determine the spontaneous lethal mutation frequency in the region balanced by szT1 gave results different from those obtained with eT1. McKim, K.S., Howell, A.M., and Rose, A.M. (1988) detected unusual segregants among the progeny of a strain of genotype dpy-5 unc-13/szT1[lon-2] I; unc-3/szT1 X. This strain has a wild-type phenotype and normally segregates wild types, Dpy-5 Unc-13 Unc-3 homozygotes, a few Lon-2 males (as a result of meiotic nondisjunction of the X chromosome), and aneuploids that arrest as embryos or young larvae. From among the progeny of 1104 unmutagenized wild-type heterozygotes, 34 animals were recovered that did not give Dpy Unc-13 Unc-3 progeny and thus appeared to carry a new lethal in one of the balanced regions. These animals, however, did not give progeny appropriate for an szT1 heterozygote and, on further analysis, appeared to carry a compound chromosome consisting of szT1 (X) fused to the normal X carrying the unc-3 mutation. These types of spontaneous rearrangements reduce the utility of szT1 as a balancer for lethal screens, although it has been used successfully after mutagenesis to recover lethals linked to dpy-5 (McKim, K.S., Howell, A.M., and Rose, A.M., 1988).
In properly designed strains, animals heterozygous for a balancer and a balanced mutation (and marker mutations), or homozygous for the mutations and carrying a balancing duplication, possess a unique phenotype. These strains can be maintained by selecting animals with this phenotype and checking their progeny to be sure that appropriate phenotypic classes are segregated. The genetic details vary among individual balancers, and a good understanding of these details is required to maintain a given strain. Specific information on normal progeny genotypes and phenotypes for each class of balancer can be found Section 2 (Types of balancers), and information on homozygous balancer phenotypes and standard marker mutations for particular balancers can be found Section 5 (A field guide to balancers).
The original isolates of some balancers carry mutations that were in the genetic background of the strains from which the balancers were generated (e.g., dpy-10 and unc-52 in the case of mnC1) or carry mutations in genes interrupted by rearrangement breakpoints (e.g., unc-36 in eT1). In addition, existing balancers have been specially marked with lethal or morphological mutations for particular tasks. These variants can be useful for strain construction (Rosenbluth, R.E., and Baillie, D.L., 1981), maintenance of balanced mutations (Johnsen and Baillie, unpublished results), and analysis of mutations and balancer structure (Rosenbluth, R.E., and Baillie, D.L., 1981). They can be generated by different means, including spontaneous mutation (Rogalski, T.M., Bullerjahn, A.M.E., and Riddle, D.L., 1988), mutagenesis or recombination (Rosenbluth, R.E., and Baillie, D.L., 1981), and DNA fusion (Hunter, C.P., and Wood, W.B., 1990). The new mutations they carry may be recessive or dominant and may be conditional or non-conditional.
Lethal variants of homozygous viable balancers can be used when it is desirable to remove balancer homozygotes from a population, leaving heterozygotes as the only viable class of progeny. For example, Johnsen and Baillie (unpublished results) used a lethal derivative of eT1 to maintain lethal mutations in strains in which balancer homozygotes were the most vigorous progeny class. Lethal variants are useful to keep balancer homozygotes from overgrowing a population under any conditions in which heterozygotes cannot be selected routinely, such as large-scale preparations for biochemical analysis or DNA extraction.
Morphologically marked balancer variants are used when a homozygous viable balancer does not already have a unique phenotype, when its phenotype is similar to that of one of the progeny classes, or when a second scorable phenotype is needed in addition to the balancer's native homozygous phenotype. For example, Rosenbluth, R.E., and Baillie, D.L. (1981) induced new mutations on eT1 to determine the segregation patterns of the half-translocations. Zetka, M.C., and Rose, A.M. (1992) induced a new unc-54 mutation on the inversion hIn1, which originally had a wild-type homozygous phenotype, to distinguish homozygotes from heterozygotes.
Most of the lethal or morphological balancer variants in common use were generated by mutagenesis with EMS and are thus not very likely to be structurally altered. That is, they may generally be expected to balance the same regions as their parent balancers. On the other hand, balancers subjected to mutagenesis with ionizing radiation could be expected to carry new rearrangements (Rosenbluth, R.E., Cuddeford, C., and Baillie, D.L., 1985; McKim, K.S., and Rose, A.M., 1990; McKim, K.S., Peters, K., and Rose, A.M., 1993). Thus, it may be necessary to test the extent of balancing for particular variants.
Crosses to move mutations into or out of different genetic backgrounds are basic to genetic analysis. For example, crosses are necessary to genetically map mutations; to complement them against deficiencies, duplications, or other mutations; to study the effects of different mutations on each other; and to remove unwanted deleterious mutations after mutagenesis. If one wishes to cross a balanced mutation into another genetic background, males that carry the mutation and are able to mate may be needed. Three methods can be employed to accomplish this. First, a mixed population of heterozygous males, half of which carry the balancer and half of which carry the balanced mutation, can be made by crossing heterozygous hermaphrodites with wild-type males. Second, males can arise in balanced strains by spontaneous X-chromosome nondisjunction, the frequency of which can be increased by a him mutation in the genetic background (Hodgkin, J.A., Horvitz, H.R., and Brenner, S., 1979) or by heat stock (Sulston, J., and Hodgkin, J., 1988), and these males can be used to propagate balanced male stocks. Third, and most desirable, balanced phenotypically unique males can be made from each balanced strain on demand by crossing to a standard male balancer strain.
The first method complicates analysis by requiring F2 progeny testing to ensure that the correct marker has been transferred. The second method mayor may not require progeny testing, depending on whether the desired F2 cross-progeny have a unique phenotype; however, it requires that a male stock be maintained for each balanced mutation. Strain construction and complementation tests are made much simpler if the genotypes and corresponding phenotypes resulting from crosses are unique, and if the required males can be generated as needed.
The following discussion presents a few examples of useful crossing protocols of the latter class for particular balancers. The original published works, or the investigators themselves, are still the best source for specific detail and advice.
1. hIn1 (I) Ho and Rose (unpublished results) devised a simple scheme to facilitate the complementation testing of a set of lethal mutations induced using the inversion hIn1 as a balancer. A single cross for each balanced lethal generated wild-type males of a single genotype that could be used in complementation tests against deficiencies and other lethal mutations. For example, males of a standard balancer strain, of genotype hIn1[unc-54]/unc-75 unc-101, were crossed to hIn1[unc-54 unc-75]/unc-101 lev-11 let-x hermaphrodites (Figure 5). The only phenotypically wild-type male progeny, of genotype hIn1[unc-54]/unc-101 lev-11 let-x, were then crossed to other identically balanced lethals in complementation tests (e.g., hIn1[unc-54]/unc-101 lev-11 let-y). In this case, complementation of let-x by let-y was indicated by the presence of viable, fertile Unc-101 Lev-11 progeny (unc-101 lev-11 let-x/unc-101 lev-11 let-y). Failure to complement was indicated by the absence of this progeny class. The same scheme was used to map the lethals against deficiencies that deleted both lev-11 and unc-54 (such as eDf3). In these cases, complementation or its failure was indicated by the presence or absence of Lev-11 progeny, as none of the deficiencies deleted unc-101.
|
Figure 5. Example of a crossing scheme using hIn1 to generate genotypically and phenotypically unique balanced males carrying a desired lethal mutation. The standard male balancer strain and a sample balanced lethal strain are represented by genotypes in the boxes on the left. The genotypes and phenotypes of the four classes of cross progeny resulting are shown on the right. Both male and hermaphrodite cross progeny will be present. This example illustrates a basic principle of marker manipulation, which can be adapted for other balancers and markers.
2. szT1(I;X) An advantage of szT1 in crossing protocols is its spontaneous segregation of fertile males that have a distinct Lon-2 phenotype and are hemizygous for the translocation. These males have been used effectively for strain construction and complementation tests (McKim, K.S., and Rose, A.M., 1990) and elucidation of the structure of the translocation (McKim, K.S., Howell, A.M., and Rose, A.M., 1988). The translocation was induced in a strain carrying lon-2(e678), and this mutation is thus present on one half-translocation (Fodor, A., and Deak, P., 1985). Males from szT1 strains carrying lon-2 must have a Lon-2 phenotype, as they are hemizygous for the X chromosome. These males carry both half-translocations and one copy of a normal LG I. As males arise spontaneously in szT1 strains at a frequency of approximately 10% (Fodor, A., and Deak, P., 1985; McKim, K.S., and Rose, A.M., 1990), it is not necessary to make males that carry either the half-translocations or the balanced LG I markers by crossing balanced hermaphrodites with wild-type males.
The three basic uses of the Lon males depend on their exclusive transfer of the normal LG I to all viable male progeny and of the translocation chromosomes to all viable hermaphrodite progeny (Figure 6). First, the males can be used to balance existing mutations. For example, dpy-5 could be balanced with szT1 simply by crossing Lon-2 males to a dpy-5 homozygote. As all wild-type hermaphrodite progeny resulting from the cross must be heterozygous for both szT1 and dpy-5, the mutation is balanced in a single step. Second, the males can be used to generate wild-type males heterozygous for szT1-balanced lethal mutations for use in complementation tests against other szT1-balanced lethals, assuming the lethals have a morphological marker in common and assuming that failure to complement can be scored reliably in male progeny. Third, they can be used for purposes that require F3 hermaphrodite progeny carrying two copies of the normal LG I. For example, McKim, K.S., and Rose, A.M. (1990) used unc-11 dpy-14/szT1 (I); szT1 (X)/O males to generate wild-type males of genotype unc-11 dpy-14/dpy-5 let-x unc-13. These were then crossed to other strains to test for complementation of let-x by a set of duplications. Presence of the unc-11 dpy-14 chromosome allowed them to select progeny of the desired genotype without progeny testing.
3. mnDp1 (X;V) This duplication of part of the right end of LG X is translocated to LG V. It is sterile as a homozygote, but heterozygous males are fertile (Herman, R.K., Albertson, D.G., and Brenner, S., 1976). Normally, analysis of X-linked mutations is complicated because males are hemizygous for the X chromosome and thus express the mutant phenotype. For example, males resulting from crossing wild-type males to a homozygous mutant unc-3 (LG X) hermaphrodite would have the genotype unc-3/0 (X). These would have an Unc-3 phenotype and be unable to mate or mate poorly. The use of mnDp1 (and similar duplications for other parts of the X chromosome) solves this problem. A hermaphrodite strain carrying two mutant copies of unc-3 and one copy of mnDp1 has a wild-type phenotype, as the duplication provides a wild-type copy of the unc-3 gene. Males resulting from crossing wild-type males with this strain have one of two genotypes and correspondingly unique phenotypes. Half will have the genotype +/+ V; unc-3/O and be Unc-3, and half will have the genotype mnDp1/+ V; unc-3/0 and a wild-type phenotype. The latter are able to mate and transfer both the duplication and the unc-3 mutation in crosses. Meneely and Herman (1979) used these properties of mnDp1 to analyze recessive lethal mutations balanced by mnDp1 that are either linked to or delete unc-3. Wild-type males of genotype mnDp1/+ V; unc-3 let-x/O were used to transfer the two X mutations into different genetic backgrounds for two-and three-factor mapping and for complementation tests with other balanced lethals.
|
Figure 6. Example of a crossing scheme using spontaneous Lon-2 males from an szT1 strain to balance a dpy-5 mutation. The male and hermaphrodite parents are represented by genotypes in the boxes on the left. The genotypes and phenotypes of the two classes of viable cross-progeny are shown on the right. Male parents are hemizygous for LG X and, because of the lon-2 mutation carried on szT1, must have a Lon phenotype. All wild-type male progeny will be heterozygous for dpy-5 and will not carry the translocation. All wild-type hermaphrodite progeny will be heterozygous for both dpy-5 and the translocation. All other cross-progeny are aneuploid, and arrest during development.
Rosenbluth, R.E., Cuddeford, C., and Baillie, D.L. (1983) measured the spontaneous lethal mutation frequency in the 40-mu region balanced by eT1 and found it to be 0.06%. Extrapolated to the whole genome, approximately 1 in every 119 selfed hermaphrodites may be expected to be heterozygous for a new lethal mutation. These mutations would quickly be lost by segregation if they appeared in strains that carried no balancer and were maintained in culture. If, however, the mutations occur in a balanced region, segregational and recombinational loss will be eliminated. Indeed, spontaneous mutations of all types (not only lethal) tend to accumulate in balancer strains. Great care must therefore be taken by the investigator to confirm that a given balanced strain has not acquired any additional mutations subsequent to the original mutagenesis. It is preferable to keep strains frozen if they are not actually being used in experiments, and strains that have been passaged should be checked closely. The effects of an additional lethal mutation in a strain that already carries a lethal can be especially subtle. For example, if the original mutation results in arrest at a mid-larval developmental stage, an additional mutation that causes the animal to arrest slightly earlier may not be noticed. New mutations causing later developmental arrest are phenotypically undetectable in a lethal that arrests as an embryo. These unrecognized secondary mutations may confound analyses.
The value of a balancer is reduced if balanced mutations spontaneously become unbalanced, or if balancer strains appear to acquire new mutations at high frequency. The point at which a balancer is no longer considered a balancer is when it cannot be used successfully for its intended purpose. This is the decision of the particular investigator. Although it is possible for any balancer to undergo recombination that results in loss of balanced mutations, it is important to distinguish between exceptional progeny that result from recombination and those that may result from chromosome nondisjunction, as described below. The determination of the nature of a putative breakdown event must always be documented through recovery and analysis of diagnostic progeny.
Exceptional segregants may lead the investigator to believe that recombination has occurred in a balancer strain. For example, the normal progeny of a strain carrying dpy-5 I and unc-3 X mutations balanced by szT1 (dpy-5/szT1[lon-2] I; unc-3/szT1 X) are wild-type heterozygotes, Dpy-5 Unc-3 homozygotes, Lon-2 males, lethal szT1 homozygotes, and lethal aneuploid progeny. Dpy non-Unc and Unc non-Dpy progeny are not usually produced because the markers are pseudolinked. Rare Unc-3 progeny do arise (McKim, K.S., and Rose, A.M., 1990), which at first glance could be the product of a recombination event between the normal LG I and szT1(X) that resulted in loss of dpy-5 from the normal LG I. Careful analysis of one such event, however, revealed that no recombination had occurred. Instead, one of the half-translocations, szT1(X), carrying a portion of LG I, was present as a duplication in addition to two normal copies of LG I (marked with dpy-5) and two normal copies of LG X (marked with unc-3). As this duplication, named szDp1, carried a wild-type copy of dpy-5 but not of unc-3, the strain had an Unc-3 phenotype. Analogous exceptional segregants have been recovered from a number of translocation strains, for example, mnDp11 from mnT2 (Herman, R.K., Kari, C.K., and Hartman, P.S., 1982), hDp133 from hT1, hDp134 from hT2, and hDp135 from hT3 (McKim, K.S., Peters, K., and Rose, A.M., 1993). Thus, the investigator must be aware of the possibility that a translocation heterozygote will give rise to apparent recombinant progeny and must analyze the exceptional progeny before concluding that a crossover has occurred. Physical breakdown of a few balancers has been documented. For example, some duplications of LG I have been observed to shorten spontaneously, resulting in exposure of previously balanced mutations (McKim, K.S., and Rose, A.M., 1990). Also, Zetka, M.C., and Rose, A.M. (1992) documented two rare recombination events in hIn1 heterozygotes that resulted in deficiencies. In all these cases, the fact that physical rearrangement had occured was documented through the recovery and analysis of exceptional progeny.
Balancers have been used in mutant screens most commonly for recovery of lethal mutations, but they can also be used to recover nonlethal mutations. For example, they can be used in noncomplementation screens for new alleles of existing morphological mutations. The work summarized below is concerned mostly with lethal mutations, but it provides practical information for anyone contemplating a mutant screen. The following information will not substitute for a thorough knowledge of balancer genetics and the published works, but the points are good guides for avoiding unnecessary work and confusion.
Ideally, nonbalancer strains are mutagenized and the new mutations are captured in crosses to balancers, to prevent inducing mutations on the balancer itself. Such crosses also replace approximately half the mutagenized genome with unmutagenized chromosomes, helping to remove unwanted second-site mutations. For large screens, however, routine backcrossing is not practical, and it is much more efficient to mutagenize balancer strains directly. Genetic mapping and inter se complementation of the new mutations can be accomplished rapidly with strains that are already balanced and appropriately marked, but that have not been backcrossed, as discussed previously. The problem of second-site mutations can be minimized by the use of low mutagen doses (see below). Before more extensive characterization is undertaken for particular mutations, back-crossing can be done using schemes similar to those presented in Section 3 (Practical considerations of balancer use).
Screens should be done at low mutagen doses for all mutagens to decrease the incidence of multiple mutational events. Studies undertaken by Rosenbluth, R.E., Cuddeford, C., and Baillie, D.L. (1983; 1985) demonstrated the utility of low doses of EMS and gamma irradiation, and characterized a number of rearrangements generated in these experiments. Based on these studies, we recommend doses in the range 1500 to 3000 R for gamma irradiation and 6 to 18 mM for EMS. It is tempting to use higher doses to increase the efficiency of recovery, but the benefits are more than outweighed by the increased complication of analysis. At the very least, the use of lower doses can help keep the amount of work down by reducing the need for extensive backcrossing prior to analysis. Rare multiple events are usually spotted in the process of mapping, although this is not always the case (Anderson, P., 1995).
Mutagenesis of balancer strains has yielded point mutations, deficiencies, duplications, and translocations. Some of these can be very complicated to analyze. The investigator should be conversant with all the published work involving a prospective balancer before using it to recover lethals, and should also be conversant with the detailed study by Rosenbluth, R.E., Cuddeford, C., and Baillie, D.L. (1985) of the types and frequency of mutations recovered in screens with eT1.
Screens using reciprocal translocations will recover lethals in the two genomic regions balanced by the two half-translocations. Figuring out on which chromosome they reside requires linkage tests or mapping (e.g., with deficiencies). Linkage mapping is fairly trivial if the original balanced strain carries morphological markers on both balanced chromosomes. For example, refer to Figure 2. Mapping to chromosome can be done by outcrossing to remove the translocation and examining segregating progeny for reduced numbers of either Dpy-18 (indicating a lethal on LG III) or Unc-46 (indicating a lethal on LG V). Subsequent higher-resolution positioning can be accomplished using recombination mapping and complementation tests with deficiencies or duplications.
Translocation and inversion screens can yield lethal mutations that are balanced because they are close to the rearrangement breakpoints, but they lie outside the rearrangement and may be lost through recombination. Duplications have a precise recovery boundary, as they balance by complementation of the lethal (presence of a wild-type allele). A duplication screen will only yield lethals that lie inside the genetic extents of the duplication. Deficiencies spanning the duplication endpoint will not be recovered if they delete any essential genes inside the endpoint.
The presence of morphological mutations linked to lethals in balancer strains has several benefits. First among these is the ease of recovery of the lethals. It much easier to screen for the absence of a particular morphological mutant class than it is to screen for arrested embryos or larvae. The second major advantage is that presence of the markers eases genetic mapping, complementation analysis, and arrest-stage determination of the lethals. A disadvantage is that the investigator may encounter marker effects. The presence of particular markers may result in a lethal arresting at a different developmental stage than it would without the markers, or a given mutation may be lethal in the presence of a marker but completely viable in its absence. For example, dpy-14 exhibits a synthetic lethal phenotype in doubly mutant constructs involving a number of muscle mutants, such as unc-15 and unc-54 (Rose, A.M., and Baillie, D.L., 1980). It is best remove the markers prior to phenotypic analysis.
Care must be taken when selecting homozygous progeny for further developmental or molecular characterization to understand the correspondence of phenopes and genotypes segregated from a balanced lethal heterozygote. For example, translocation strains segregate a large proportion of aneuploid progeny (Figure 2) that arrest as embryos or early larvae, but these are not the lethal homozygotes of interest. To be certain that the arrested embryos or animals are of the correct genotype, the lethal-bearing chromosome must be crossed to a nonbalanced genetic background and resegregated as a homozygote (Johnsen, R.C., and Baillie, D.L., 1991).
The advantages of balancers for lethal mutant screens seem ideal for extension to screens conducted in mutator genetic backgrounds that exhibit a high frequency of Tc1 transposition (Moerman, D.G., and Waterston, R.H., 1989; Anderson, P., 1995). The ease of maintenance and crossing is especially attractive given the necessity of removing the mutator from the background once a mutation has been induced. Although these screens may produce desired mutations, they may not be as generally useful as are screens using other mutagens. Using mut-4(st7000)I, Clark, D.V., Johnsen, R.C., McKim, K.S., and Baillie, D.L. (1990) analyzed 28 spontaneous lethal mutations (probably Tc1-induced) in the 49-mu region balanced by nT1. The distribution of these mutations was skewed: 86% of them mapped to LG V, as compared with 43% of EMS-induced lethal mutations recovered over nT1. Furthermore, the types of mutations obtained were different from those obtained using EMS, formaldehyde, or gamma irradiation. Two were new mutations in previously unidentified genes, six were mutations in lin-40, and seven were deficiencies. All of the latter deleted the left end of the chromosome, and five of them had right endpoints in or near lin-40.
The distribution of transposon-induced mutations is changing with identification methods that do not require that an insertion result in a scorable phenotype (Plasterk, R.H.A., 1995). Also, analysis of secondary deletion mutants derived from strains carrying transposon insertions is resulting in information about the null phenotypes of these genes. Balancers may prove useful in such schemes for maintaining heterozygosity of induced mutations and identifying animals homozygous for the insertions or their spontaneous derivatives.
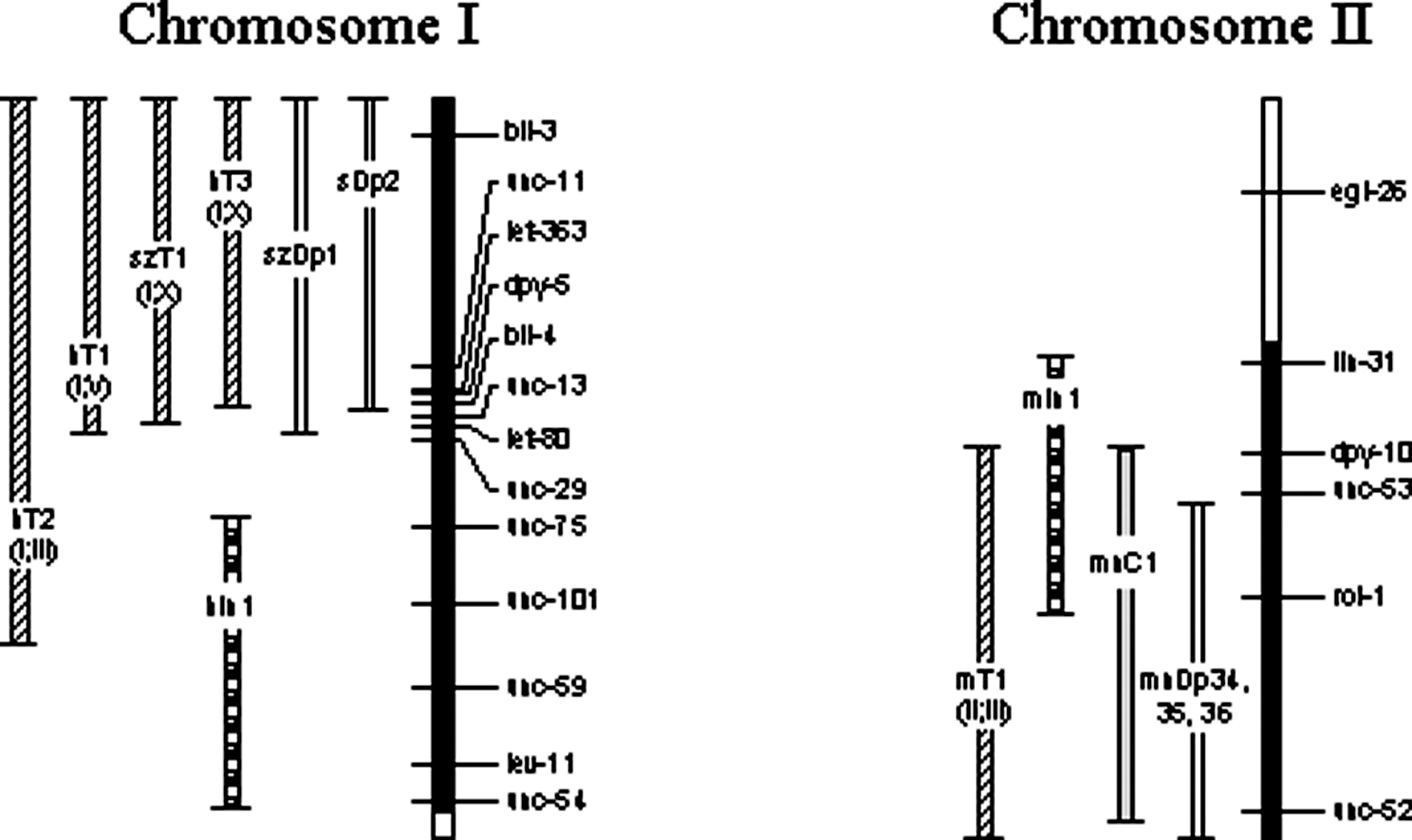
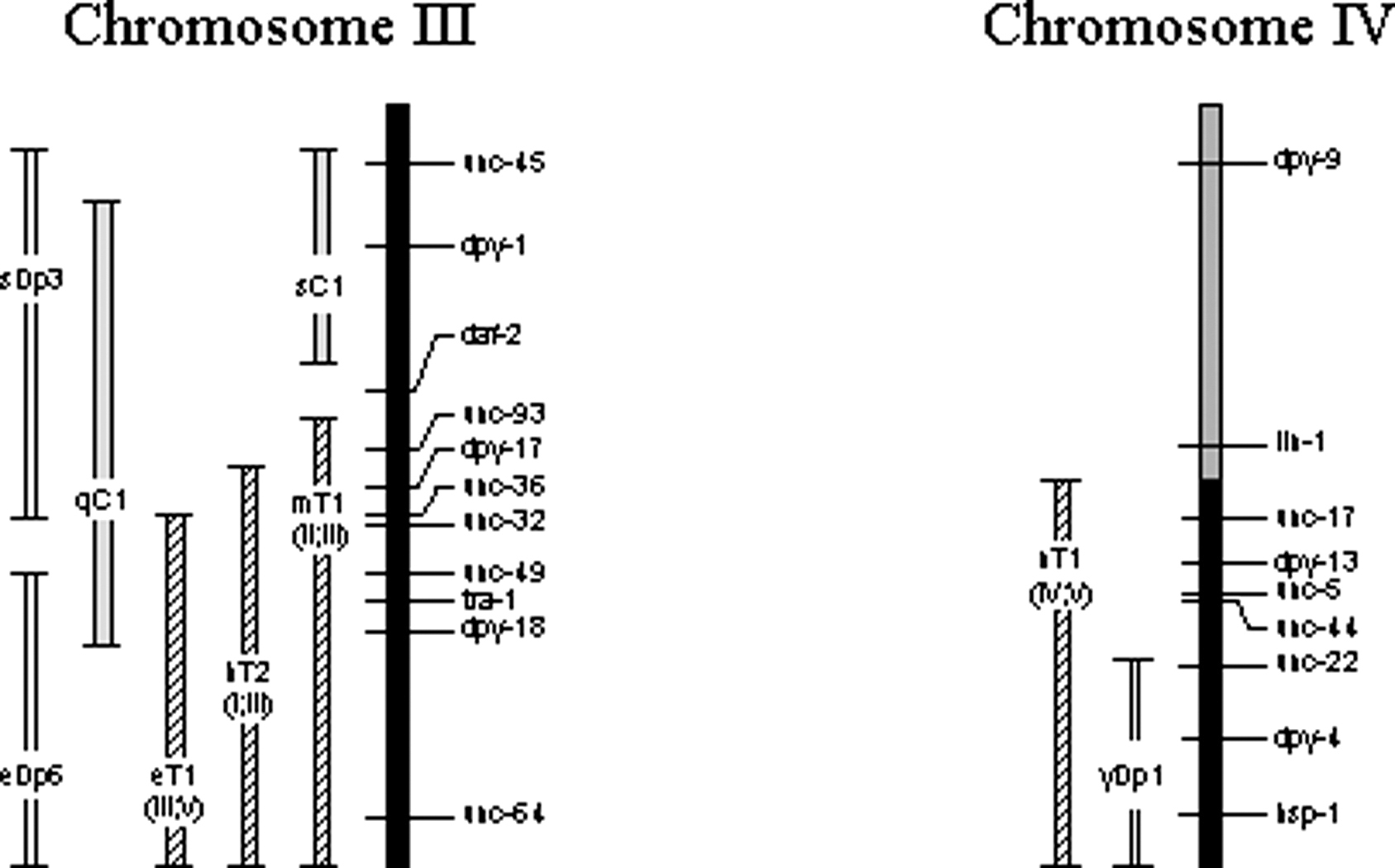
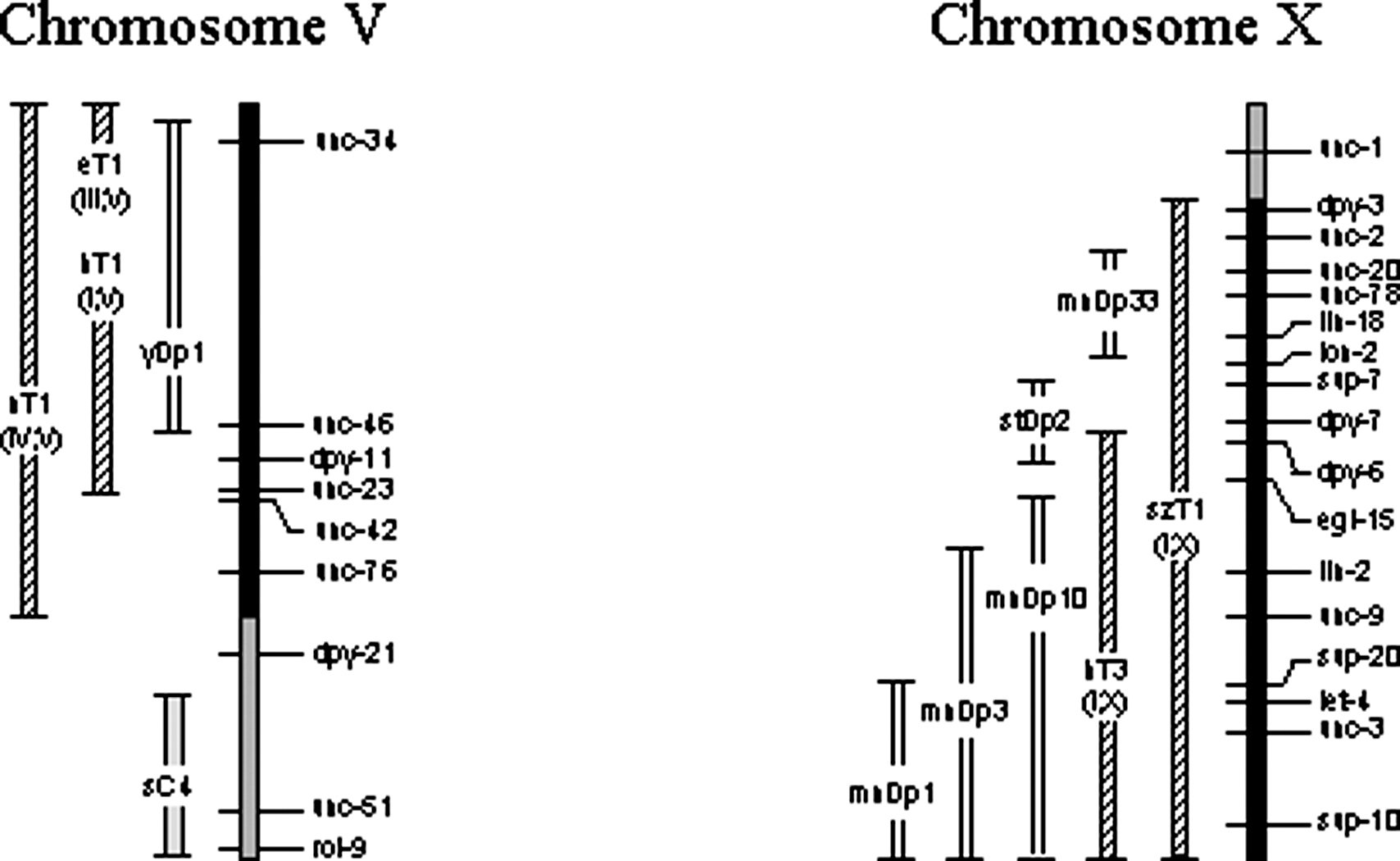
Over half the C. elegans genome is covered by reliable, tested balancers (Figure 7). Well-balanced regions include virtually all of LG I, the right 65% of LG II, all of LG III, the right half of LG IV, the left 66% of LG V, and the right 80% of LG X (based on genetic, not physical, extents of each chromosome). Incompletely balanced regions include most of the remaining areas. Balancers may or may not include the physical ends of chromosomes. When drawn or referred to as including the end, it means only that the most distal genetic marker appears to be balanced.
|
|
|
Figure 7. Map of the balanced regions of C. elegans chromosomes. Each chromosome is represented by a vertical bar with cross lines indicating genetic positions of a subset of identified genes (some genes defining the endpoints of minor balancers are not shown where space is limited). By convention, the top end of each vertically drawn chromosome corresponds to the left end when drawn horizontally, and the bottom of each corresponds to the right end. Shading of the chromosomes indicates the degree of balancing. Black represents well-balanced regions, crosshatching represents regions balanced by unstable or uncharacterized rearrangements, and no shading represents regions for which no balancers exist. Approximately 70% of the genome is well balanced. Balancers are drawn to the left of each chromosome as double-line bars (duplications) or bars with different shading patterns (other balancers). Endpoint lines on each balancer indicate its genetic extents. Some rearrangements that provide balancing of limited reliability are not shown. Also not shown are duplications that balance small intervals.
Certain chromosomal regions not covered by translocations have proven somewhat resistant to efforts to obtain effective balancers. For example, several balancers for the right arm of LG V have been recovered, but all to date have exhibited unacceptable levels of crossing over and thus are of limited value for lethal screens (Stewart and Baillie, unpublished results). Such incomplete balancers may still provide a useful degree of balancing for existing mutations, and their use should be evaluated case by case.
This guide presents alphabetically the most fully characterized and commonly used C. elegans balancers. It is by no means an exhaustive list. The "field guide" format is an attempt to present information critical for the use of each balancer in a brief and consistent manner. Standardized descriptors help compare and contrast the choices for a particular genetic region.
Most of the balancers listed here, and some of the derivatives and marked variants, are available from the Caenorhabditis Genetics Center (CGC). Those variants not stocked by the CGC generally have not been the subject of wide interest, but they may prove useful for certain screens or strain constructions. For more information about whether a particular variant is available, the investigator should contact the originating laboratory.
Most of the descriptors are self-explanatory, but a few require more detail. For translocations, inversions, and other crossover suppressors, stability refers generally to the frequency at which unusual progeny may be segregated. Measures of stability are necessarily subjective, as very little data has been accumulated on the frequency of such progeny and the nature of the events that produced them. The stability descriptor therefore represents a consensus opinion based on published information and the experiences of the authors' laboratories. Extremely stable means that neither recombination nor unusual segregants have been documented. Very stable means that rare unusual segregants have been recovered, but the utility of the balancer is not generally affected. Stable means that unusual segregants have been recovered at somewhat higher frequency, and utility of the balancer is potentially compromised. Details are given where the nature of unusual segregants has been deduced through further analysis.
For all balancers, Recommended use is broken down into four categories, presented in increasing stringency of characteristics required in the balancer. General balancing means that it is useful for small-scale use. Strain construction is listed only for balancers known to be especially useful in crossing protocols because of specific characteristics or the existence of marked variants. By definition, males carrying these balancers must be able to mate. Strain maintenance means that a balancer is stable enough for large-scale use, although it may be unsuitable for mutant screens. Mutant screens means that a balancer has been proven reliable in screens for lethal mutations.
Summary: Free duplication, moderately well characterized, not observed to recombine with normal homologs. Very effective balancer for right portion of LG III from right end through tra-1 and vab-7.
Origin: Acetaldehyde mutagenesis.
Recommended use: General balancing, strain maintenance.
Reference strain: CB1517, eDf2 III; eDp6(III;f).
Phenotype: Unc-119.
Segregants: Unc-119.
Handling: Unc phenotype of reference strain is characteristic of the combination of two copies of eDf2 and one copy of eDp6. This is due to deletion of the unc-119 locus by eDf2 and incomplete complementation of the deletion by the duplication (D. Pilgrim and M. Maduro, pers. comm.).
References: Hodgkin, J. (1980; 1987); Hunter, C.P., and Wood, W.B., 1990.
Summary: Reciprocal translocation, well characterized, extremely stable. Very effective balancer for left portion of LG V from left end through unc-23, and right portion of LG III from right end to unc-36. eT1(III) is LG V (left) translocated to LG III (left), disjoins from normal LG III. eT1(V) is LG III (right) translocated to LG V (right), disjoins from normal LG V.
Origin: 32P mutagenesis of N2.
Recommended use: General balancing, strain construction, strain maintenance, mutant screens.
Reference strain: BC2200, dpy-18(e364)/eT1 III; unc-46(e177)/eT1 V.
Phenotype: Wild type.
Segregants: Wild type, Unc-36 eT1 homozygotes, Dpy-18 Unc-46, and large numbers of arrested aneuploid progeny.
Growth characteristics: Original isolate homozygous viable with Unc-36 phenotype caused by the translocation breakpoint. Brood size in heterozygotes ~ 100, in homozygotes ~160. eT1 homozygotes can overgrow a population, especially in balanced strains carrying certain lethal mutations or deficiencies.
Handling: Easy to manipulate. Heterozygous male stocks mate well.
Marked variants: Morphological: eT1[bli-5(s277)]/eT1 [dpy-11(s287)], eT1[sma-2(s262)], eT1[sma-3(e491)], eT1[unc-42(e270)]. Lethal: eT1[let-500(s2165)].
References: Rosenbluth, R.E., and Baillie, D.L., 1981; Rosenbluth, R.E., Cuddeford, C., and Baillie, D.L. (1983; 1985); McKim, K.S., Howell, A.M., and Rose, A.M., 1988; Nelson, G.A., Schubert, W.W., Marshall, T.M., Benton, E.R., and Benton, E.V., 1989; Adames, K.A., Gawne, J., Wicky, C., Muller, F., and Rose, A.M., 1998.
Summary: Inversion, well characterized, very stable. Very effective balancer for right portion of LG I from unc-75 through unc-54. Extreme right end of chromosome apparently not balanced.
Origin: Gamma irradiation of N2 males.
Recommended use: General balancing, strain construction, strain maintenance, mutant screens.
Reference strain: KR2267, hIn1[unc-54(h1040)]/unc-75(e950) unc-101(m1) I.
Phenotype: Wild type.
Segregants: Wild type, Unc-54 hIn1 homozygotes, Unc-75 Unc-101.
Growth characteristics: Original isolate homozygous viable with wild-type phenotype; unc-54 mutation in the reference strain was induced secondarily. Brood size similar to N2 as homozygote and as heterozygote.
Handling: Easy to manipulate. Heterozygous males, and homozygous males of original isolate, mate well. Very rare recombination can occur, which generates deficiencies and duplications.
Marked variants: Morphological: hIn1[unc-54(h1040)], hIn1[unc-101(sy241)]. Lethal: hIn1[unc-75(h1041)], hIn1[unc-75(h1042)], hIn1[unc-54(h1040) unc-75(h1041)].
Derivatives: hDf11, hDf12, hDp131, hDp132.
References: Lee, J., Jongeward, G.D., and Sternberg, P.W., 1994; Zetka, M.C., and Rose, A.M., 1992.
Summary: Reciprocal translocation, well characterized, very stable. Very effective balancer for left portion of LG I from the left end through let-80, and the left portion of LG V from the left end through dpy-11. hT1(I) is LG V (left) translocated to LG I (right), disjoins from normal LG I. hT1(V) is LG I (left) translocated to LG V (right), disjoins from normal LG V.
Origin: Gamma irradiation of unc-23 + / + unc-42 males.
Recommended use: General balancing, strain maintenance, mutant screens.
Reference strain: KR1037, unc-13(e51)/hT1 I; dpy-11(e224)/hT1[unc-42(e270)] V.
Phenotype: Wild type.
Segregants: Wild type, arrested hT1 homozygotes, Unc-13 Dpy-11, and large numbers of arrested aneuploid progeny. Recombination occurs occasionally between the hT1 breakpoint and unc-42 on hT1(V), giving rise to Unc-42 animals. These are of genotype unc-13(e51)/hT1 I; dpy-11(e224) unc-42(e270)/hT1[unc-42(e270)]V.
Growth characteristics: Homozygous inviable, cause unknown. Arrests at L3. Brood size in heterozygotes ~75.
Handling: Easy to manipulate. Heterozygous males mate well. Rare exceptional progeny carry one half-translocation as a complex free duplication. Recombination frequency in the unbalanced unc-101 - unc-54 interval on LG I is increased twofold.
Marked variants: hT1[unc-29(e403)].
Derivatives: hDp133(I;V;f).
References: McKim, K.S., Howell, A.M., and Rose, A.M., 1988; Howell, A.M., and Rose, A.M., 1990.
Summary: Reciprocal translocation, well characterized, stable. Effective balancer for left portion of LG I from left end through unc-101, and right portion of LG III from right end through dpy-17. hT2(I) is LG III (right) translocated to LG I (right), disjoins from normal LG I. hT2(III) is LG I (left) translocated to LG III (left), disjoins from normal LG III.
Origin: Gamma irradiation of bli-4(e937)I males.
Recommended use: General balancing, strain maintenance.
Reference strain: KR1235, unc-13(e51)/hT2 I; dpy-18(e364)/hT2[bli-4(e937)] III.
Phenotype: Wild type.
Segregants: Wild type, Bli-4 hT2 homozygotes, Unc-13 Dpy-18, and large numbers of arrested aneuploid progeny.
Growth characteristics: Original isolate homozygous viable with Bli-4 phenotype.
Handling: Easy to manipulate. Heterozygous males, and homozygous males of original isolate, mate well. Homozygous Bli-4 phenotype completely suppressed in dpy-5 and dpy-18 variants. Rare exceptional progeny carry one half-translocation as a complex free duplication. Mutations have been observed to become unbalanced at low frequency, but the mechanism is not fully understood. The dpy-18(h662) variant is very mildly Dpy as a homozygote. hT2[bli-4(e937) let-?(q782) qIs48] carries an integrated pharyngeal GFP element; the lethal mutation in this variant has been observed to recombine away, leaving hT2[bli-4(e937) qIs48] that apparently retains balancer activity.
Marked variants: hT2[dpy-5(h659)], hT2[dpy-18(h662)], hT2[unc-54(e190)], hT2[bli-4(e937) unc-29(h1011)], hT2[dpy-18(h662) unc-59(e261)], hT2[bli-4(e937) let-?(q782) qIs48].
Derivative: hDp134 (I;III;f).
References: McKim, K.S., Peters, K., and Rose, A.M., 1993; J. Kimble, pers. comm.
Summary: Translocation (rigorous proof of reciprocity lacking), moderately well characterized, very stable. Very effective balancer for left portion of LG I from left end to around let-363, and the right portion of LG X from the right end to between dpy-7 and unc-3. hT3(I), which disjoins from normal LG I, is probably LG X (right) translocated to LG I (right). hT3(X),which disjoins from normal LG X, is probably LG I (left) translocated to LG X (left).
Origin: Gamma irradiation mutagenesis of dpy-5(e61) unc-29(e403)/hT2[dpy-18(h662)] I; +/hT2[bli-4(e937)] III. Isolated as a lethal mutation balanced by hT2, now maintained as a homozygote balanced by szDp1.
Recommended use: General balancing, strain maintenance.
Reference strain: KR1879, hT3[dpy-5(e61) unc-29(e403)](I;X); szDp1(I;X;f).
Phenotype: Unc-29 (szDp1 complements dpy-5 but not unc-29).
Segregants: Unc-29, arrested hT3 homozygotes.
Growth characteristics: Original isolate marked with dpy-5 and unc-29. Homozygous inviable, probably breaks in let-363 (I). Heterozygotes exhibit reduced viability, low level of X chromosome nondisjunction (1.2%).
Handling: Easy to manipulate. Rare exceptional progeny carry one half-translocation as a free duplication. Recombination frequency in unbalanced intervals increased on both LG I and LG X.
Marked variants: hT3[dpy-5(e61)].
Derivative: hDp135(I;X;f).
Reference: McKim, K.S., Peters, K., and Rose, A.M., 1993.
Summary: Inversion, well characterized, very stable. Very effective balancer for center portion of LG II from lin-31 through rol-1.
Origin: Gamma irradiation of dpy-10(e128)/unc-85(e1414) rol-1(e91)II.
Recommended use: General balancing, strain construction, strain maintenance, mutant screens.
Reference strain: DR1785, mIn1[dpy-10(e128)]/unc-4(e120)II.
Phenotype: Wild type.
Segregants: Wild type, Dpy-10 mIn1 homozygotes, Unc-4.
Growth characteristics: Original isolate marked with dpy-10(e128). Brood size similar to N2 as heterozygote. Homozygotes do not survive freezing well.
Handling: Easy to manipulate. Heterozygous males mate well. mIn1[dpy-10(e128) mIs14] carries an integrated pharyngeal GFP element; expression is semi-dominant, such that one copy of mIs14 can be distinguished from two by GFP signal brightness. Rearrangement may have occurred in the integration event, as lethal mutations in the far left extent of the balanced region are not as stable over the GFP variant as they are over the original mIn1.
Marked variants: Morphological: mIn1[unc-4(e120) dpy-10(e128)], mIn1(+), mIn1[unc-4(e120)], mIn1[rol-1(e91)], mIn1[rol-1(e91) dpy-10(e128)], mIn1[dpy-10(e128) mIs14(myo-2::GFP)]. Lethal: mIn1[dpy-10(e128) let-?(m727)].
Reference: Edgley, M.L., and Riddle, D.L., 2001.
Summary: Reciprocal translocation, well characterized, very stable. Very effective balancer for right portion of LG II from the right end through dpy-10, and the right portion of LG III from the right end to between daf-2 and unc-93. mT1(II) is probably LGII (right) translocated to LG III (left), disjoins from normal LG III. mT1(III) is probably LG III (right) translocated to LG II (left), disjoins from normal LG II.
Origin: Gamma irradiation of dpy-10(e128)/unc-85(e1414) rol-1(e91) II.
Recommended use: General balancing, strain maintenance, mutant screens.
Reference strain: DR1832, mT1/unc-4(e120)II; mT1[dpy-10(e128)]/dpy-17(e164)III.
Phenotype: Wild type.
Segregants: Wild type, sterile Dpy mT1 homozygotes, Unc-4 Dpy-17, and large numbers of arrested aneuploid progeny.
Growth characteristics: Original isolate marked with dpy-10(e128). Translocation probably breaks in an essential gene, as mT1 homozygotes are sterile.
Handling: Easy to manipulate. Heterozygous males mate reasonably well.
Reference: Edgley, M.L., and Riddle, D.L., 2001.
Summary: Dominant crossover suppressor, uncharacterized with regard to structure, very stable. Very effective balancer for right portion of LG II from around dpy-10 to around unc-52.
Origin: X-ray mutagenesis of unc-4/dpy-10 unc-52.
Recommended use: General balancing, strain construction, strain maintenance, mutant screens.
Reference strain: SP127, unc-4(e120)/mnC1[dpy-10(e128) unc-52(e444)].
Phenotype: Wild type.
Segregants: Wild type, Unc-4, Dpy-10 Unc-52 mnC1 homozygotes. The homozygotes are short, fat, and paralyzed, whereas the Unc-4 segregants are large and healthy and move forward well, but cannot back up.
Growth characteristics: Homozygotes viable with extremely small broods (average: 7 progeny). Brood size in heterozygotes ~200, with 100% egg hatching.
Handling: Easy to manipulate. Heterozygous male stocks mate well. Very rare recombination gives rise to Dpy non-Unc and Unc non-Dpy progeny. The recombinant chromosomes carried by these progeny are homozygous lethal.
References: Herman, R.K., 1978; Sigurdson, D.C., Spanier, G.J., and Herman, R.K., 1984.
Summary: Translocated duplication, well characterized, does not recombine with normal homologues. Very effective balancer for right portion of LG X from right end through let-4.
Origin: ~7500-R X-ray mutagenesis of N2 males.
Recommended use: General balancing, strain construction, strain maintenance, mutant screens.
Reference strain: SP219, mnDp1(X;V)/+ V; unc-3(e151)X.
Phenotype: Wild type.
Segregants: Wild type, Unc-3, sterile mnDp1 homozygotes.
Growth characteristics: Brood size of reference strain ~235. Unc-3 progeny constitute 25% of total, and about a third of total are slow-growing, sterile duplication homozygotes.
Handling: Easy to manipulate. Males mate well. mnDp1 is attached to LG V near the left end, and severely reduces recombination frequency from unc-60 to dpy-11.
References: Herman, R.K., Albertson, D.G., and Brenner, S., 1976; Herman, R.K., Madl, J.E., and Kari, C.K., 1979; Meneely P.M., and Herman, R.K., 1981.
Summary: Free duplication, well characterized, does not recombine with normal LG X. Effective balancer for right portion of LG X from around the right end through unc-9.
Origin: ~7500-R X-ray mutagenesis of N2 males.
Recommended use: General balancing, strain construction, strain maintenance.
Reference strain: SP123, unc-3(e151)X; mnDp3(X;f).
Phenotype: Wild type.
Segregants: Wild type, Unc-3.
Growth characteristics: Brood size ~275, duplication transmitted to ~65% of progeny.
Handling: Easy to manipulate. Males carrying duplication mate well. Shows some tendency for somatic loss. Marked variant: mnDp3[sup-10(mn338)].
References: Herman, R.K., Albertson, D.G., and Brenner, S., 1976; Herman, R.K., Madl, J.E., and Kari, C.K., 1979; Herman, R.K., 1984.
Summary: Translocated duplication, well characterized, does not recombine with normal LG X. Effective balancer for right portion of LG X from around the right end through lin-2.
Origin: ~7500-R X-ray mutagenesis of N2 males.
Recommended use: General balancing, strain construction, strain maintenance.
Reference strain: SP117, mnDp10(X;I); unc-3(e151)X.
Phenotype: Wild type.
Segregants: Wild type, rare Unc-3.
Growth characteristics: Duplication is homozygous viable. Unc-3 animals arise from somatic loss of duplication.
Handling: Easy to manipulate. Males carrying the duplication mate well. mnDp10 shows some tendency to segregate from the X chromosome in male spermatogenesis.
References: Herman, R.K., Madl, J.E., and Kari, C.K., 1979; Meneely P.M., and Herman, R.K., 1981.
Summary: Translocated duplication, well characterized, does not recombine with normal X. Effective balancer for small region of left portion of LG X, from lin-18 to osm-5 (inclusive).
Origin: ~7500 R X-ray mutagenesis of N2 males.
Recommended use: General balancing; strain construction, strain maintenance.
Reference strain: SP309, mnDp33(X;IV); unc-20(e112)X.
Phenotype: Wild type.
Segregants: Wild type, Unc-20, arrested L1 or L2 larvae (duplication homozygotes).
Growth characteristics: Homozygous inviable. ~100% egg hatching, but 25% arrest.
Handling: Easy to manipulate. Males carrying the duplication mate well.
Reference: Herman, R.K., Madl, J.E., and Kari, C.K., 1979.
Summary: Independently isolated free duplications with apparently identical genetic extents. Well characterized, do not recombine with normal homologues. Very effective balancers for right portion of LG II from around unc-52 through unc-53.
Origin: ~7500-R X-ray mutagenesis of mnC1[dpy-10(e128) unc-52(e444)]/ unc-4(e120).
Recommended use: General balancing, strain maintenance.
Reference strains:
| mnDp34: SP306, mnC1[dpy-10 unc-52]/unc-4 unc-52; mnDp34. |
| mnDp35: SP307, mnC1[dpy-10 unc-52]/unc-4 unc-52; mnDp35. |
| mnDp36: SP308, mnC1[dpy-10 unc-52]/unc-4 unc-52; mnDp36. |
Phenotype: Wild type.
Segregants: Wild type, Dpy-l0 Unc-52 (mnC1[dpy-10 unc-52] homozygotes), Unc-52 (unc-4 unc-52 homozygotes and mnC1[dpy-10 unc-52]/unc-4 unc-52), Unc-4 (unc-4 unc-52; mnDpx), Dpy-l0 (mnC1[dpy-10 unc-52]; mnDpx).
Growth characteristics: Egg hatching nearly 100%. About 20% of these arrest development as early larvae, and are presumably duplication homozygotes. The duplications are transmitted to about 40% of progeny.
Handling: These duplications tend to segregate from the X chromosome in male spermatogenesis.
Reference: Herman, R.K., Madl, J.E., and Kari, C.K., 1979.
Summary: Translocation, moderately well characterized, very stable. Very effective balancer for right portion of LG IV from right end through unc-17, and for left portion of LG V from left end through unc-76.
Origin: Spontaneous.
Recommended use: General balancing, strain maintenance, mutant screens.
Reference strain: MTl000, unc-5(e53)/nTl IV; dpy-11(e224)/nTl V.
Phenotype: Wild type.
Segregants: Wild type, vulvaless nT1 homozygotes that become bags of worms, Unc-5 Dpy-11, and large numbers of arrested aneuploid progeny.
Growth characteristics: Brood size in nT1 heterozygotes ~100.
Handling: Easy to manipulate. Heterozygous males mate well. Vulvaless homozygous hermaphrodites completely unable to mate. Cause of vulvaless phenotype unknown. Translocation may break down spontaneously, but analysis of such events is lacking. Heterozygous strains occasionally begin to segregate large numbers of sick-looking progeny while appearing to remain heterozygous, or they occasionally begin to give larger broods (Schein and Baillie, unpublished results). nT1[qIs51] carries an integrated pharyngeal GFP element, and is homozygous inviable; this variant appears to be transferred to male cross progeny more frequently than to hermaphrodite cross progeny.
Marked variants: nT1[unc(n754dm) let] (previously called DnT1, dominant Unc, recessive Let); nT1[qIs51]; nTl[let(m435)].
References: Ferguson, E.L., and Horvitz, H.R., 1985; Clark, D.V., Rogalski, T.M., Donati, L.M., and Baillie, D.L., 1988; Rogalski, T.M., and Riddle, D.L., 1988; J. Kimble, pers. comm.
Summary: Dominant crossover suppressor, uncharacterized with regard to structure, very stable. Very effective balancer for the left portion of LG III from tra-1 to at least dpy-1.
Origin: 7200-R gamma-irradiation mutagenesis of unc-32(e189)/dpy-19(e1259).
Recommended use: General balancing, strain maintenance.
Reference strain: JK1122, dpy-17(e164) unc-32(e189)/qC1[dpy-19(e1259) glp-1(q339)] III.
Phenotype: Wild type.
Segregants: Wild type, Dpy-17 Unc-32 homozygotes, sterile ts Dpy-19.
Handling: Relatively easy to manipulate. Heterozygous males mate and transfer qC1, though perhaps at low frequency. qC1 fails to complement glp-1 and mog-1 mutations.
Marked variants: qC1[dpy-19(e1259) glp-1(q339) qIs26].
References: Austin, J., and Kimble, J., 1989; J. Austin, E. Goodwin, and J. Kimble, pers. comm.; Ha and Baillie, unpublished results.
Summary: Dominant crossover suppressor, uncharacterized with regard to structure, very stable. Very effective balancer for an approximately 15-mu portion of LG III from around unc-45 to near daf-2.
Origin: 2000-R gamma-irradiation mutagenesis of N2 males.
Recommended use: General balancing, strain maintenance, mutant screens.
Reference strain: BC4279, sC1[dpy-l (s2170)] III.
Phenotype: Dpy-l.
Segregants: Dpy-l (homozygous strain).
Growth characteristics: Original isolate wild type. Broods approximately wild type in size.
Handling: Easy to manipulate. Heterozygous males, and homozygous males of original isolate, mate well.
Marked variants: sC1[dpy-1(s2171)]; sC1[dpy-1(s2171) let].
Reference: Stewart and Baillie, unpublished results.
Summary: Dominant crossover suppressor, uncharacterized with regard to structure, moderately stable. Balances right portion of LG V from rol-9 to unc-76.
Origin: 2000-R gamma-irradiation mutagenesis of dpy-21/unc-76 rol-9.
Recommended use: General balancing.
Reference strain: BC4586, sC4[dpy-21]/unc-76 rol-9.
Phenotype: Wild type.
Segregants: Wild type, Unc-76 Rol-9, arrested sC4 homozygotes. Presence of dpy-21 on sC4 not confirmed.
Growth characteristics: Homozygous inviable. Average brood size 117. Cause of homozygous lethal phenotype unknown. unc-76 - rol-9 genetic distance reduced to 1.8%.
Handling: Easy to manipulate. Heterozygous males mate well.
Reference: Stewart and Baillie, unpublished results.
Summary: Free duplication, well characterized, does not recombine with normal homologues. Very effective balancer for the left portion of LG I from the left end through unc-15 (just left of unc-13).
Origin: 7500-R gamma-irradiation mutagenesis of N2 males.
Recommended use: General balancing, strain maintenance, mutant screens.
Reference strain: KR236, dpy-5(e61) unc-13(e450)I; sDp2(I,f).
Phenotype: Unc-13.
Segregants: Unc-13, Dpy-5 Unc-13.
Growth characteristics: Both Unc-13 and Dpy-5 Unc-13 animals are slow growing (generation time at 20°C nearly 5 days). Animals carrying two copies of sDp2 have never been recovered.
Handling: sDp2-bearing males mate and give some progeny, but are slow growing and do not compete well with non-Dp males in mating.
Derivatives: hDp2-hDp30; hDp59; hDp74-hDp77.
References: Rose, A.M., Baillie, D.L., and Curran, J., 1984; Howell, A.M., Gilmour, S.G., Mancebo, R.A., and Rose, A.M., 1987; Howell, A.M., and Rose, A.M., 1990; McKim, K.S., and Rose, A.M., 1990; McKim, K.S., Starr, T., and Rose, A.M., 1992; McKim, K.S., Peters, K., and Rose, A.M., 1993.
Summary: Free duplication, well characterized, does not recombine with normal homologues. Very effective balancer for left portion of LG III from around unc-86 through at least dpy-1 (does not extend to unc-45).
Origin: 1500-R gamma-irradiation mutagenesis of dpy-18/eT1 III; unc-46/ eT1 V.
Recommended use: General balancing, strain maintenance, mutant screens.
Reference strain: BC986, eT1(III;V); sDp3(III,f).
Phenotype: Wild type.
Segregants: Wild type, Unc-36 eT1 homozygotes.
Handling: Easy to manipulate. Males mate well. Duplication homozygotes are probably inviable (never recovered).
Reference: Rosenbluth, R.E., Cuddeford, C., and Baillie, D.L., 1985.
Summary: Translocated duplication, moderately well characterized, not observed to recombine with normal LG X. Very effective balancer for small region in the center of LG X, from around unc-58 to around unc-6.
Recommended use: General balancing, strain construction, strain maintenance.
Reference strain: RW6002, +/stDp2 II; unc-18(e81)X.
Phenotype: Wild type.
Segregants: Wild type, Unc-18.
Growth characteristics: Homozygous inviable.
Handling: Easy to manipulate. Males carrying one copy of the duplication mate well.
Reference: Meneely, P.M. and Wood, W.B., 1984.
Summary: Complex free duplication, well characterized, does not recombine with normal LG I. Very effective balancer for the left portion of LG I from the left end through unc-13. Consists of one half-translocation [szT1(X)] from szT1 maintained in addition to a normal chromosome complement.
Origin: Exceptional Unc-3 segregant from dpy-5/szT1[lon-2] I; unc-3/szT1 X.
Reference strain: KRI577, dpy-5(e61) unc-13(e450)I; szDpI(I;x,f).
Phenotype: Wild type.
Segregants: Wild-type hermaphrodites, small percentage wild-type males, Dpy-5 Unc-13.
Growth characteristics: Animals carrying two copies of szDp1 apparently inviable. Duplication strains give rise to spontaneous males through meiotic nondisjunction of the X chromosome.
Handling: szDp1-bearing males either do not mate or are infertile.
Derivatives: hDp31-hDp58; hDp60-hDp73.
References: McKim, K.S., Howell, A.M., and Rose, A.M., 1988; McKim, K.S., Starr, T., and Rose, A.M., 1992; McKim, K.S., Peters, K., and Rose, A.M., 1993; McKim, K.S., and Rose, A.M., 1990.
Summary: Reciprocal translocation, well characterized, very stable. Effective balancer for left portion of LG I through unc-13, nearly all of LG X from right end to around dpy-3. szT1(I) is large segment of LG X (right) translocated to LG I, disjoins from normal LG I. szT1(X) is LG I (left) translocated to fragment of LG X (left), disjoins from normal LG X.
Origin: 7000-R gamma-ray mutagenesis of lon-2(e678)/dpy-8(e1321) unc-3(e151)hermaphrodites.
Recommended use: General balancing, strain construction, strain maintenance.
Reference strain: AFl, +/szT1[lon-2(e678) I; dpy-8(e1321) unc-3(e151)/szT1 X.
Phenotype: Wild type.
Segregants: Wild type, Dpy-8 Unc-3 hermaphrodites and males, embryonic lethal szT1 homozygotes, large numbers of embryonic or early larval arrest aneuploid progeny, and about 10% Lon-2 males.
Growth characteristics: Homozygous inviable; breakpoint resulting in lethality must interrupt a gene on LG I, as hemizygous males are viable and fertile. Lon-2 males arise through meiotic nondisjunction of X chromosome. Brood size in heterozygotes ~100.
Handling: Easy to manipulate. Lon-2 szT1 males mate well. Rare exceptional progeny carry one half-translocation as a complex free duplication. Gives rise spontaneously to rare apparent lethal mutations that may represent fusion of szT1(X) and the normal X. Shows threefold enhanced recombination frequency immediately adjacent to right of LG I breakpoint and about twofold enhanced frequency in the unc-101 - unc-54 interval.
Marked variant: szT1[lon-2(e678) unc-29(e403)].
References: Fodor, A., and Deak, P., 1985; McKim, K.S., Howell, A.M., and Rose, A.M., 1988; Howell, A.M., and Rose, A.M., 1990; McKim, K.S., and Rose, A.M., 1990.
Summary: Complex free duplication, moderately well characterized, not observed to recombine with either normal homologue. Very effective balancer for right portion of LG IV from around dpy-4 to around unc-22, and left portion of LG V from around unc-34 through unc-46.
Origin: EMS mutagenesis of unc-22(s7) dpy-26(n199)IV/nT1[unc(n754) let](IV;V).
Recommended use: General balancing, strain construction, strain maintenance.
Reference strain: TY156, unc-30(e191) dpy-4(e1166) IV; yDp1(IV;V,f).
Phenotype: Wild type.
Segregants: Wild type, Unc-30 Dpy-4.
Growth characteristics: Brood size ~250. Animals carrying two copies of yDp1 apparently inviable. The duplication segregates from X chromosome in male meiosis.
Handling: Easy to manipulate. Males carrying the duplication mate well.
References: DeLong, L., Casson, L.P., and Meyer, B.J., 1987; Plenefisch, J.D., DeLong, L., and Meyer, B.J., 1989; Yuan, J.Y., and Horvitz, H.R., 1990.
Adames, K.A., Gawne, J., Wicky, C., Muller, F., and Rose, A.M. (1998). Mapping a telomere using the translocation eT1(III;V) in Caenorhabditis elegans. Genetics 150, 1059–1066. Abstract
Albertson, D.G., Fishpool, R.M., and Birchall, P.S. (1995). Fluorescence in situ hybridization for the detection of DNA and RNA. In: Methods in Cell Biology, Vol. 48, H.F. Epstein and D.C. Shakes, eds. (San Diego, CA: Academic Press), pp. 339–364. Abstract
Albertson, D.G., and Thomson, J.N. (1982). The kinetochores of Caenorhabditis elegans. Chromosoma 86, 409–428. Abstract Article
Albertson, D.G., and Thomson, J.N. (1993). Segregation of holocentric chromosomes at meiosis in the nematode, Caenorhabditis elegans. Chromosome Res. 1, 15–26. Abstract Article
Anderson, P. (1995). Mutagenesis. In: Methods in Cell Biology, Vol. 48, H.F. Epstein and D.C. Shakes, eds. (San Diego, CA: Academic Press), pp. 31–54. Abstract
Austin, J., and Kimble, J. (1989). Transcript analysis of glp-l and lin-12, homologous genes required for cell interactions during development of C. elegans. Cell 58, 565–571. Abstract Article
Brenner, S. (1974). The genetics of Caenorhabditis elegans. Genetics 77, 71–94. Abstract
Bucher, E.A., and Greenwald, I. (1991). A genetic mosaic screen of essential zygotic genes in Caenorhabditis elegans. Genetics 128, 281–292. Abstract
C. elegans Sequencing Consortium (1998). Genome sequence of the nematode C. elegans: a platform for investigating biology. Science 282, 2012–2018. Abstract Article
Chalfie M., Tu, Y., Euskirchen, G., Ward W.W., and Prasher, D.C. (1994). Green fluorescent protein as a marker for gene expression. Science 263, 802–805 Abstract
Clark, D.V., Johnsen, R.C., McKim, K.S., and Baillie, D.L. (1990). Analysis of lethal mutations induced in a mutator strain that activates transposable elements in Caenorhabditis elegans. Genome 33, 109–114. Abstract
Clark, D.V., Rogalski, T.M., Donati, L.M., and Baillie, D.L. (1988). The unc-22(IV) region of Caenorhabditis elegans: genetic analysis of lethal mutations. Genetics 119, 345–353. Abstract
DeLong, L., Casson, L.P., and Meyer, B.J. (1987). Assessment of X-chromosome dosage compensation in Caenorhabditis elegans by phenotypic analysis of lin-l4. Genetics 117, 657–670. Abstract
Edgley, M.L., and Riddle, D.L. (2001). LG II balancer chromosomes in Caenorhabditis elegans: mT1(II;III) and the mIn1 set of dominantly and recessively marked inversions. Mol. Genet. Genomics 266(3), 385–395. Abstract Article
Ferguson, E.L., and Horvitz, H.R. (1985). Identification and characterization of 22 genes that affect the vulval cell lineages of the nematode Caenorhabditis elegans. Genetics 110, 17–72. Abstract
Fodor, A., and Deak, P. (1985). The isolation and genetic analysis of a Caenorhabditis elegans translocation (szT1) strain bearing an X-chromosome balancer. J. Genet. 64, 143–157. Abstract
Greenwald, I.S., and Horvitz, H.R. (1980). unc-93(e1500): a behavioral mutant of Caenorhabditis elegans that defines a gene with a wild-type null phenotype. Genetics 96, 147–164. Abstract
Greenwald, I.S., Sternberg, P.W., and Horvitz, H.R. (1983). The lin-12 locus specifies cell fates in C. elegans. Cell 34, 435–444. Abstract Article
Herman, R.K. (1978). Crossover suppressors and balanced recessive lethals in Caenorhabditis elegans. Genetics 88, 49–65. Abstract
Herman, R.K. (1984). Analysis of genetic mosaics of the nematode Caenorhabditis elegans. Genetics 108, 165–180. Abstract
Herman, R.K. (1987). Mosaic analysis of two genes that affect nervous system structure in Caenorhabditis elegans. Genetics 116, 377–388. Abstract
Herman, R.K. (1995). Mosaic analysis. In: Methods in Cell Biology, Vol. 48, H.F. Epstein and D.C. Shakes, eds. (San Diego, CA: Academic Press), pp. 123–143. Abstract
Herman, R.K., Albertson, D.G., and Brenner, S. (1976). Chromosome rearrangements in Caenorhabditis elegans. Genetics 83, 91–105. Abstract
Herman, R.K., and Kari, C.K. (1989). Recombination between small X chromosome duplications and the X chromosome in Caenorhabditis elegans. Genetics 121, 723–737. Abstract
Herman, R.K., Kari, C.K., and Hartman, P.S. (1982). Dominant X-chromosome nondisjunction mutants of Caenorhabditis elegans. Genetics 102, 379–400. Abstract
Herman, R.K., Madl, J.E., and Kari, C.K. (1979). Duplications in Caenorhabditis elegans. Genetics 92, 419–435. Abstract
Hodgkin, J. (1980). More sex-determination mutants of Caenorhabditis elegans. Genetics 96, 649–664. Abstract
Hodgkin, J. (1987). A genetic analysis of the sex-determining gene, tra-1, in the nematode Caenorhabditis elegans. Genes Dev. 1, 731–745. Abstract
Hodgkin, J.A., Horvitz, H.R., and Brenner, S. (1979). Nondisjunction mutants of the nematode Caenorhabditis elegans. Genetics 91, 67–94.
Horvitz, H.R., Brenner, S., Hodgkin, J., and Herman, R.K. (1979). A uniform genetic nomenclature for the nematode Caenorhabditis elegans. Mol. Gen. Genet. 175, 129–133. Abstract Article
Howell, A.M., Gilmour, S.G., Mancebo, R.A., and Rose, A.M. (1987). Genetic analysis of a large autosomal region in Caenorhabditis elegans by the use of a free duplication. Genet. Res. 49, 207–213.
Howell, A.M., and Rose, A.M. (1990). Essential genes in the hDf6 region of chromosome I in Caenorhabditis elegans. Genetics 126, 583–592. Abstract
Hunter, C.P., and Wood, W.B. (1990). The tra-1 gene determines sexual phenotype cell-autonomously in C. elegans. Cell 63, 1193–1204. Abstract Article
Johnsen, R.C., and Baillie, D.L. (1988). Formaldehyde mutagenesis of the eTl balanced region in .. Caenorhabditis elegans: dose-response curve and the analysis of mutational events. Mutat. Res. 201, 137–147.
Johnsen, R.C., and Baillie, D.L. (1991). Genetic analysis of a major segment [LGV(left)] of the genome of Caenorhabditis elegans. Genetics 129, 735–752. Abstract
Lee, J., Jongeward, G.D., and Sternberg, P.W. (1994). unc-101, a gene required for many aspects of Caenorhabditis elegans development and behavior, encodes a clathrin-associated protein. Genes Dev. 8, 60–73. Abstract
Howell, A.M., and Kose, A.M. (1988). The effects of translocations on recombination frequency in Caenorhabditis elegans. Genetics 120, 987–1001.
Marra, M.A., and Baillie, D.L. (1994). Recovery of duplications by drug resistance selection in Caenorhabditis elegans. Genome 37, 701–705. Abstract
McKim, K.S., Howell, A.M., and Rose, A.M. (1988). The effects of translocations on recombination frequency in Caenorhabditis elegans. Genetics 120, 987–1001. Abstract
McKim, K.S., and Rose, A.M. (1990). Chromosome I duplications in Caenorhabditis elegans. Genetics 124, 115–132. Abstract
McKim, K.S., Peters, K., and Rose, A.M. (1993). Two types of sites required for meiotic chromosome pairing in Caenorhabditis elegans. Genetics 134, 749–768. Abstract
McKim, K.S., Starr, T, and Rose, A.M. (1992). Genetic and molecular analysis of the dpy-14 region in Caenorhabditis elegans. Mol. Gen. Genet. 233, 241–25l. Abstract Article
Mello, C., and Fire, A. (1995). DNA transformation. In: Methods in Cell Biology, Vol. 48, H.F. Epstein and D.C. Shakes, eds. (San Diego, CA: Academic Press), pp. 452–480. Abstract
Mello, C.C., Kramer, J.M., Stinchcomb, D., and Ambros, V. (1991). Efficient gene transfer in C. elegans: extrachromosomal maintenance and integration of transforming sequences. EMBO J. 10, 3959–3970. Abstract
Meneely, P.M., and Herman, R.K. (1979). Lethals, steriles and deficiencies in a region of the X chromosome of Caenorhabditis elegans. Genetics 92, 99–115. Abstract
Meneely, P.M., and Herman, R.K. (1981). Suppression and function of X-linked lethal and sterile mutations in Caenorhabditis elegans. Genetics 97, 65–84. Abstract
Meneely, P.M., and Nordstrom, K.D. (1988). X chromosome duplications affect a region of the chromosome they do not duplicate in Caenorhabditis elegans. Genetics 119, 365–375. Abstract
Meneely, P.M., and Wood, W.B. (1984). An autosomal gene that affects X chromosome expression and sex determination in Caenorhabditis elegans. Genetics 106, 29–44. Abstract
Moerman, D.G., and Baillie, D.L. (1979). Genetic organization in Caenorhabditis elegans: fine-structure analysis of the unc-22 gene. Genetics 91, 95–104.
Moerman, D.G., and Baillie, D.L. (1981). Formaldehyde mutagenesis in the nematode Caenorhabditis elegans. Mutat. Res. 80, 273–279. Abstract
Moerman, D.G., and Waterston, R.H. (1989). Mobile elements in Caenorhabditis elegans and other nematodes. In Mobile DNA, D.E. Berg and M.M. Howe, eds. (Washington, D. C: American Society for Microbiology), pp. 537–556.
Nelson, G.A., Schubert, W.W., Marshall, T.M., Benton, E.R., and Benton, E.V. (1989). Radiation effects in Caenorhabditis elegans, mutagenesis by high and low LET ionizing radiation. Mutat. Res. 212, 181–192. Abstract
Plasterk, R.H.A. (1995). Reverse genetics: From gene sequence to mutant worm. In: Methods in Cell Biology, Vol. 48, H.F. Epstein and D.C. Shakes, eds. (San Diego, CA: Academic Press), pp. 59–78. Abstract
Plenefisch, J.D., DeLong, L., and Meyer, B.J. (1989). Genes that implement the hermaphrodite mode of dosage compensation in Caenorhabditis elegans. Genetics 121, 57–76. Abstract
Riddle, D.L., and Brenner, S. (1978). Indirect suppression in Caenorhabditis elegans. Genetics 89, 299–314. Abstract
Rogalski, T.M., Bullerjahn, A.M.E., and Riddle, D.L. (1988). Lethal and amanitin-resistance mutations in the Caenorhabditis elegans ama-1 and ama-2 genes. Genetics 120, 409–422. Abstract
Rogalski, T.M., and Riddle, D.L. (1988). A Caenorhabditis elegans RNA polymerase II gene, ama-I IV, and nearby essential genes. Genetics 118, 61–74. Abstract
Rose, A.M., and Baillie, D.L. (1980). Genetic organization of the region around unc-15(I), a gene affecting paramyosin in Caenorhabditis elegans. Genetics 96, 639–648. Abstract
Rose, A.M., Baillie, D.L., and Curran, J. (1984). Meiotic pairing behavior of two free duplications of linkage group I in Caenorhabditis elegans. Mol. Gen. Genet. 195, 52–56. Abstract Article
Rosenbluth, R.E., and Baillie, D.L. (1981). The genetic analysis of a reciprocal translocation, eT1(III;V), in Caenorhabditis elegans. Genetics 99, 415–428. Abstract
Rosenbluth, R.E., Cuddeford, C., and Baillie, D.L. (1983). Mutagenesis in Caenorhabditis elegans. 1. A rapid eukaryotic mutagen test system using the reciprocal translocation eTI(III;V). Mutat. Res. 110, 39–48.
Rosenbluth, R.E., Cuddeford, C, and Baillie, D.L. (1985). Mutagenesis in Caenorhabditis elegans. II. A spectrum of mutational events induced with 1500 Rofgamma-radiation. Genetics 109, 493–511. Abstract
Rosenbluth, R.E., Johnsen, R.C., and Baillie, D.L. (1990). Pairing for recombination in LG V of Caenorhabditis elegans: a model based on recombination in deficiency heterozygotes. Genetics 124, 615–625. Abstract
Rosenbluth, R.E., Rogalski, T.M., Johnsen, R.C., Addison, L.M., and Baillie, D.L. (1988). Genomic organization in Caenorhabditis elegans: deficiency mapping on linkage group V (left). Genet. Res. 52, 105–118.
Sigurdson, D.C., Spanier, G.J., and Herman, R.K. (1984). Caenorhabditis elegans deficiency mapping. Genetics 108, 331–345. Abstract
Stewart, H.I., Rosenbluth, R.E., and Baillie, D.L. (1991). Most ultraviolet irradiation induced mutations in the nematode Caenorhabditis elegans are chromosomal rearrangements. Mutat. Res. 249, 37–54. Abstract
Sulston, J., and Hodgkin, J. (1988). Methods. In The Nematode Caenorhabditis elegans, W. B. Wood, ed. (New York: Cold Spring Harbor Laboratory), pp. 592.
Yandell, M.D., Edgar, L.S., and Wood, W.B. (1994). Trimethylpsoralen induces small deletion mutations in Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 91, 1381–1385. Abstract
Yuan, J.Y., and Horvitz, H.R. (1990). The Caenorhabditis elegans genes ced-3 and ced-4 act cell autonomously to cause programmed cell death. Dev. Biol. 138, 33–41. Abstract Article
Zetka, M.-C., and Rose, A.M. (1992). The meiotic behavior of an inversion in Caenorhabditis elegans. Genetics 131, 321–332. Abstract
*Edited by David Fay. WormMethods editor, Victor Ambros. Last revised February 25, 2005. Published April 6, 2006. This chapter should be cited as: Edgley, M. L. et al. Genetic balancers (April 6, 2006), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.89.1, http://www.wormbook.org.
§Adapted from: Edgley, M., Baillie, D.L., Riddle, D.L., and Rose, A.M. (1995). Genetic Balancers. In Methods Cell Biol. aka C. elegans: Modern Biological Analysis of an Organism, H.F. Epstein and D.C. Shakes, eds. (San Diego: Academic Press), 48, 148–184.
†To whom correspondence should be addressed. E-mail: edgley@interchange.ubc.ca, baillie@sfu.ca, driddle@genomebc.ca or arose@gene.nce.ubc.ca
‡Gene names are italicized and consist of three lowercase letters, followed by a hyphen and isolation number for individual genes. The name is usually a mnemonic referring to the nature of mutations in the gene. For example, the name dpy refers to a class of mutation characterized by a short, fat body, and stands for dumpy. The name dpy-18 refers to a particular dpy gene. Some other names used throughout the chapter are let (lethal), unc (uncoordinated), lon (long), rol (roller), and bli (blistered). Phenotypes are represented by nonitalicized gene names with the first letter capitalized. For example, the general phenotype for all dpy mutations is written Dpy, and the phenotype for dpy-18 mutations is written Dpy-18. Rearrangement names are italicized and consist of a lower case laboratory allele prefix followed by a capitalized abbreviation indicating the type of rearrangement, an isolation number, and optional chromosomal origin and location information in parentheses. For example, the name sDp2(I;f) represents the second chromosomal duplication isolated in the Baillie laboratory (allele prefix "s"). The "I;f" indicates that it is a duplication of part of LG I and that it is "free" (not attached to another chromosome). For translocations, the parenthetical information indicates the chromosomes involved. For example, eT1(III;V) indicates a translocation between LG III and LG V. The rearrangement abbreviations used here are C (dominant crossover supressor), Df (deficiency), Dp (duplication), In (inversion), and T (translocation).