Embryo series courtesy of Einhard Schierenberg
Embryo series courtesy of Einhard SchierenbergAbstract
C. elegans feeding depends on the action of the pharynx, a neuromuscular pump that joins the mouth to the intestine. The pharyngeal muscle captures food—bacteria—and transports it back to the intestine. It accomplishes this through a combination of two motions, pumping and isthmus peristalsis. Pumping, the most visible and best understood of the two, is a cycle of contraction and relaxation that sucks in liquid from the surrounding environment along with suspended particles, then expels the liquid, trapping the particles. Pharyngeal muscle is capable of pumping without nervous system input, but during normal rapid feeding its timing is controlled by two pharyngeal motor neuron types. Isthmus peristalsis, a posterior moving wave of contraction of the muscle of the posterior isthmus, depends on a third motor neuron type.
Feeding motions are regulated by the presence and quality of food in the worm's environment. Some types of bacteria are better at supporting growth than others. Given a choice, worms are capable of identifying and seeking out higher-quality food. Food availability and quality also affect behavior in other ways. For instance, given all the high-quality food they can eat, worms eventually become satiated, stop eating and moving, and become quiescent.
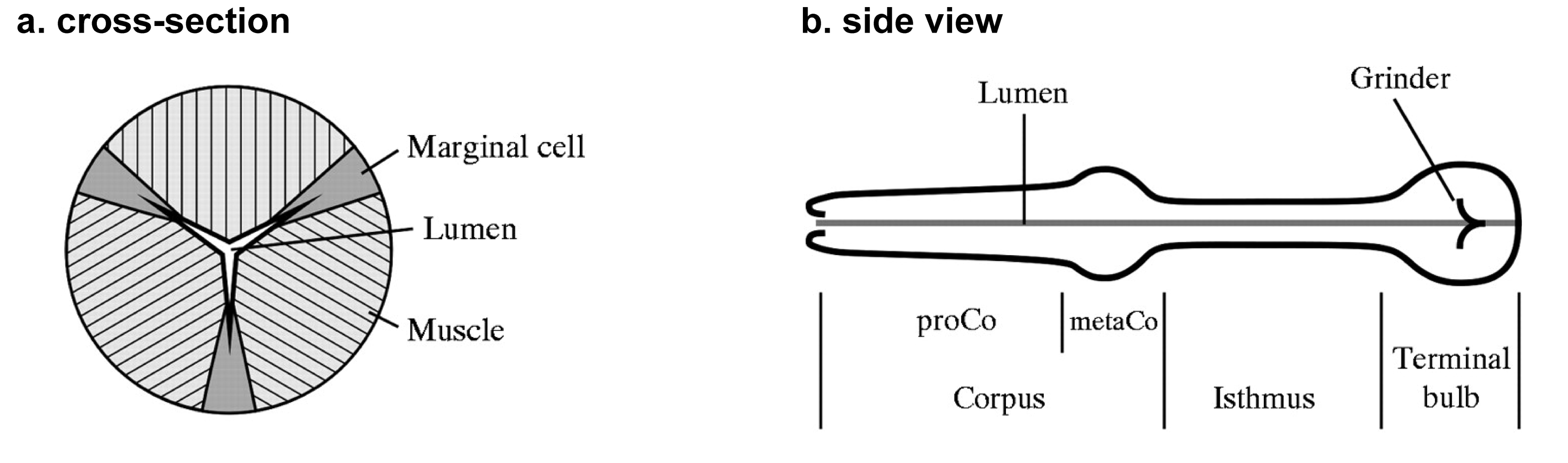
The pharynx is a tubular pump responsible for sucking bacteria into the worm from outside, concentrating them, and grinding them up (Doncaster, 1962: Seymour et al., 1983). Its cellular and nuclear composition is shown in Table 1, and the arrangement of the cells is diagrammed in Figure 1. Figure 1 and the following anatomical description are based on EM reconstructions by Albertson and Thomson (1976). This review is concerned with two of the five cell types, the muscle cells and the neurons. The pharyngeal muscle is divided into three functional groups: the corpus, the isthmus, and the terminal bulb (TB) (Figure 1b).
Table 1. Cells and nuclei in the pharynx
| Cell type | Cells | Nuclei |
|---|---|---|
| muscle | 20 | 37 |
| neurons | 20 | 20 |
| glands | 4 | 5 |
| epithelial cells | 9 | 9 |
| marginal cells | 7 | 9 |
| Total | 60 | 80 |
|
Figure 1: Anatomy of the pharynx. A. Simplified generic cross-section through the pharynx, showing the geometric relationships between muscle cells, marginal cells, and the pharyngeal lumen. B. Side view of the pharynx, showing the main functional regions. Adapted, with permission, from Avery and Horvitz (1989), and Raizen and Avery (1994).
The pharyngeal nervous system contains 20 neurons of 14 different types (8 bilaterally symmetric pairs and 6 unpaired) (Albertson and Thomson, 1976). Three of these 14 are particularly important for normal feeding. M4 is necessary and sufficient for normal isthmus peristalsis, and is essential for growth under some conditions (Avery and Horvitz, 1987). MC and M3 control the timing of pumping: MC controls when a contraction starts (Raizen and Avery, 1994; Raizen et al., 1995), and M3 when it ends (Avery, 1993; Raizen and Avery, 1994). Surprisingly, these three neuron types are sufficient for nearly normal behavior in the laboratory: even when all 11 other neuron types are killed together, the worm grows well and shows only subtle feeding abnormalities.
Pharyngeal neurons contain both neuropeptides and small-molecule neurotransmitters. The most important of the small transmitters are acetylcholine, glutamate, and serotonin. What we know of their functions is described below. Neuropeptides are certain to play important roles, but they are still for the most part poorly understood. For more information about peptides and feeding see Franks et al. (2006) and Papaioannou et al. (2005).
Pharyngeal neurons lie under the basal lamina that surrounds the entire pharynx and isolates it from the rest of the worm, in indentations of the muscle cell membrane. Aside from the connections to the mouth and the intestine there are only two holes in the basal lamina, one on either side, through which the processes of a pair of extrapharyngeal neurons, the RIPs, pass to connect to the pharyngeal nervous system.
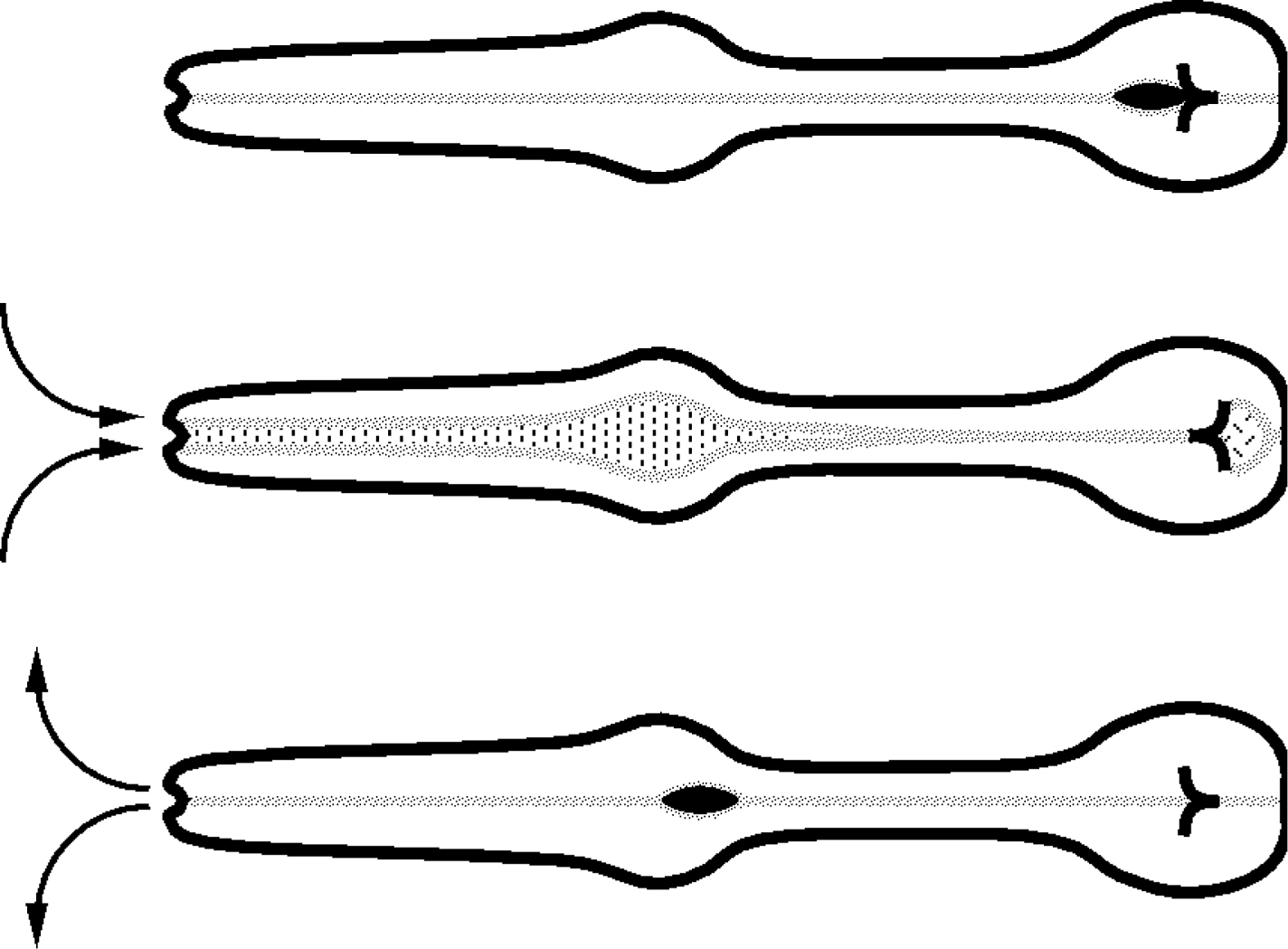
There are two feeding motions. The motion that gets food into the worm is called pumping (Figure 2). (The following description is based on Avery (1993) and references therein.) A pump begins with a near-simultaneous contraction of the muscles of the corpus, anterior isthmus, and terminal bulb. Since the pharyngeal muscles are radially oriented, contraction pulls the lumen open to a triangular cross-section, sucking in liquid and suspended bacteria from outside. Contraction is followed by a near-simultaneous relaxation, which expels liquid but traps bacteria. Correct timing of these motions is critical (Avery, 1993). The second feeding motion, isthmus peristalsis, occurs shortly after some but not all pumps. It is a peristaltic contraction of the muscles of the posterior isthmus that carries food from the corpus to the TB (Avery and Horvitz, 1987).
|
Figure 2: Pharyngeal pumping. A pharyngeal pump begins with a near-simultaneous contraction of the muscles of the corpus, anterior isthmus, and TB. Contraction of corpus and isthmus muscles pulls the lumen open. Liquid and suspended particles flow in through the mouth. At nearly the same time the TB muscles contract, inverting the plates of the grinder, which grind up bacteria and pass the debris back to the intestine. The near-simultaneous contraction is followed by a near-simultaneous relaxation that returns the grinder to its resting position and expels liquid from the corpus and isthmus. Adapted, with permission, from Avery and Horvitz (1989).
It was not at first obvious how particles are separated from liquid during pumping and transported posteriorly, since there is no obvious filter in the pharyngeal lumen. Avery and Shtonda (Avery and Shtonda, 2003) proposed that a slight difference in timing between corpus and isthmus contractions together with trapping of bacteria by the walls of the closed pharyngeal lumen could account for posteriorward transport. Fang-Yen and Avery (Fang-Yen et al., 2009), using high-speed video recordings confirmed some but not all parts of this model. In addition, they identified two filtering mechanisms that allow rough particle size-selection.
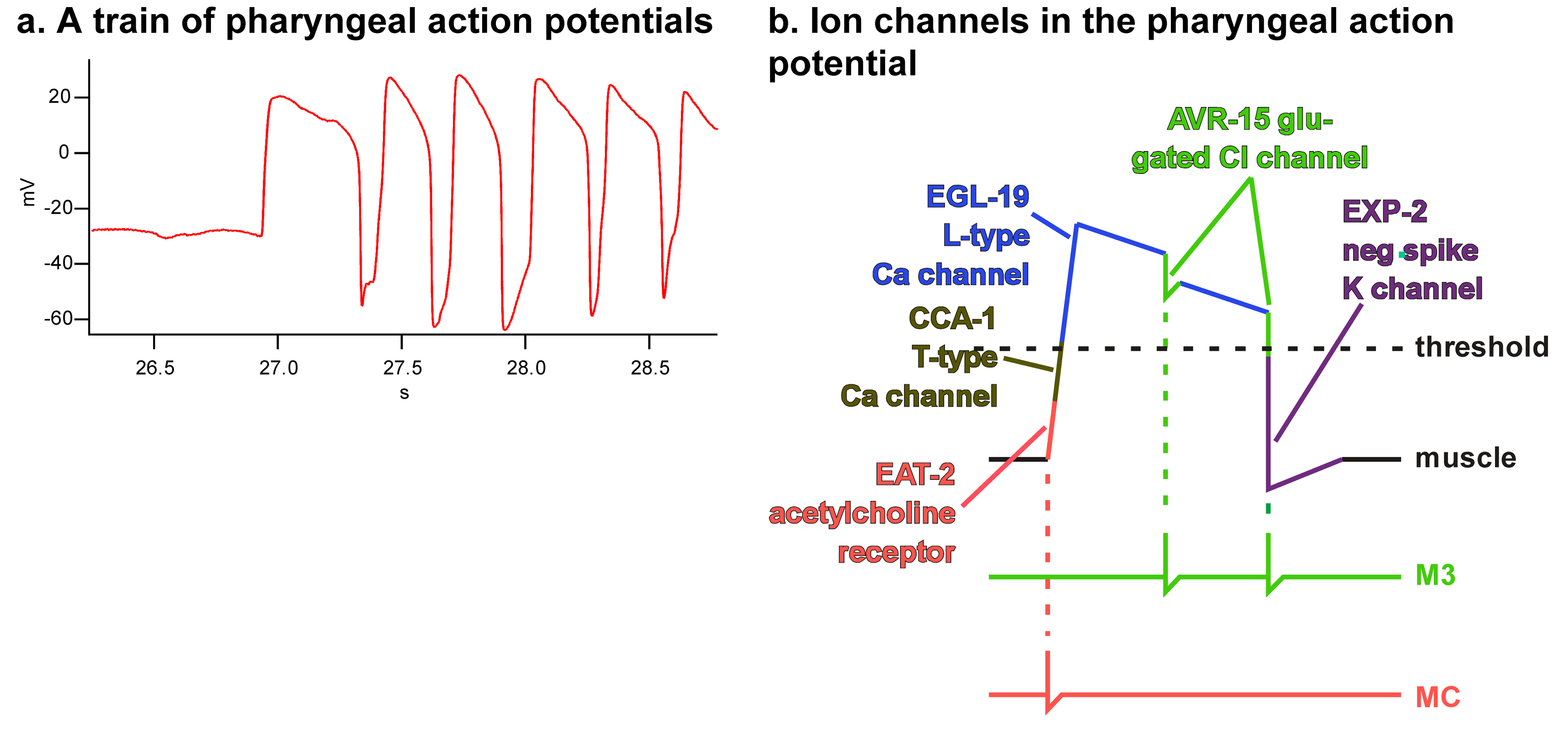
The pharynx must generate these two motions, pumping and isthmus peristalsis, at the right frequency and with the right timing. Pumping is the better understood of the two. Each contraction/relaxation cycle corresponds to a single pharyngeal muscle action potential. The action potential is produced through the sequential action of five ion channels: a nicotinic acetylcholine receptor activated by pharyngeal MC motor neurons, a low-threshold voltage-gated calcium channel, a high-threshold voltage-gated calcium channel, a glutamate-gated chloride channel activated by pharyngeal M3 motor neurons, and a voltage-gated potassium channel.
Figure 3a shows a train of action potentials recorded from TB muscle (M. Wayne Davis, personal communication). The pharynx is quiet for the first half second, then begins to pump, first with a long-lasting (400 ms) action potential, followed by a regular series of shorter (150 ms) action potentials. Besides the shape of the typical pharyngeal muscle action potential, this figure shows that action potentials, and therefore pumping, are regulated in both frequency and duration. Both neurons and muscle-intrinsic molecules contribute to this regulation.
|
Figure 3: The pharyngeal muscle action potential. A. A train of pharyngeal action potentials recorded from TB muscle. This recording was chosen to illustrate features of the action potential discussed in the text. The features visible in this recording are not always observed. In particular, the resting potential is often much more negative, and the positive slope between action potentials is not always seen (Cook et al., 2006). B. Schematic pharyngeal muscle action potential showing the action of the major ion channels.
During rapid regular pumping each action potential is initiated by the firing of MC motor neurons, whose activity can be detected indirectly by means of the postsynaptic potentials they produce in muscle (Raizen and Avery, 1994; Raizen et al., 1995). Fast neurotransmission at the MC→pharyngeal muscle neuromuscular junction (NMJ) is mediated by acetylcholine acting on nicotinic receptors (McKay et al., 2004; Raizen and Avery, 1994; Raizen et al., 1995). In the presence of food adult worms pump 200–300 times per minute on average, and eat-2 and eat-18 are necessary for this rapid pumping. eat-2 encodes a nicotinic receptor subunit, expressed in pharyngeal muscle and localized to the MC→corpus muscle NMJ (McKay et al., 2004). A functional EAT-2::GFP fusion is found only at this NMJ. eat-18 encodes a small novel single-pass transmembrane protein, also expressed in pharyngeal muscle, that is necessary for EAT-2 function, probably by direct binding to EAT-2. α-bungarotoxin (a specific nicotinic receptor probe) binds to much of the pharyngeal muscle surface. EAT-18 is necessary for this binding, but EAT-2 is not (McKay et al., 2004), suggesting there are multiple pharyngeal muscle nicotinic receptors. EAT-2 is a subunit of one of them specific for the MC NMJ, and EAT-18 is necessary for the proper targeting or function of most or all of them. Alan Chiang (personal communication) has confirmed these hypotheses by showing that, in mutants lacking the calcium-activated potassium channel SLO-1, EAT-18 but not EAT-2 is necessary for M4→TB muscle transmission.
Neither eat-2 (Raizen et al., 1995) nor MC (Avery and Horvitz, 1989) is necessary for pumping. In the absence of MC function, the pharyngeal muscle will spontaneously generate action potentials, which lead to functional pumping. Pumping in the absence of MC function is slow and irregular, and the worms grow slowly, but are nonetheless viable in the lab. (MC might of course be more important in the wild—in the lab, C. elegans is grown literally swimming in food.) This slow, irregular, MC-independent pumping may also occur in normal worms in the absence of food. It is not known how MC-independent action potentials are initiated. Based on ion substitution experiments, Franks et al. (Franks et al., 2002) proposed the existence of a hyperpolarization-induced sodium channel, functionally somewhat similar to that of vertebrate cardiac muscle, that might serve this purpose. (See also Vinogradova et al., 2006). They also suggested that a voltage-gated sodium channel contributes to the action potential. Neither of these channels has been identified molecularly.
Low threshold-activated or T-type calcium channels are postulated to mediate a variety of bursting and rhythmic electrical firing events. They allow excitable cells to respond to small depolarizations from rest, and mediate low-threshold calcium spikes (Perez-Reyes, 2003). Shtonda and Avery (2005) showed that cca-1 encodes the α1 (pore-forming) subunit of a T-type Ca2+ channel active at the beginning of the pharyngeal muscle action potential. Steger et al. (Steger et al., 2005) showed that cca-1 is necessary for efficient initiation of action potentials in response to stimulation by the MC motor neuron. When wild-type cca-1 is absent, the depolarizing phase of the pharyngeal action potential tends to plateau or stall near −30mV, the voltage at which the CCA-1 channel is likely to be activated. Therefore, loss of cca-1 function decreases the chance that excitatory input from MC will successfully trigger an action potential, and reduces the ability of an animal to take in food.
Thus, the CCA-1 T-type calcium channel boosts the excitatory effect of synaptic input, allowing for reliable and rapid depolarization and contraction of the pharyngeal muscle. However, pharyngeal muscle employs alternative strategies for initiating action potentials in certain cases of compromised MC motor neuron function as in eat-2 ((Shtonda and Avery, 2005; Steger et al., 2005).
egl-19 encodes the α1 subunit of a homolog of vertebrate L-type voltage-activated Ca2+ channels and plays a pivotal role in regulating muscle excitation and contraction in all C. elegans muscles. Raymond Lee (Lee et al., 1997) isolated several egl-19 mutants and sorted them into three classes: (1) myotonic group (semidominant, gain-of-function mutations in egl-19 cause myotonia), mutant muscle action potentials are prolonged and the relaxation delayed; (2) flaccid group, partial loss-of function mutations cause slow muscle depolarization and feeble contraction; and (3) lethal group, the most severe loss-of-function mutants (including nulls) lack all muscle contraction and die as embryos. Lee et al. (1997) localized two myotonic mutations in the sixth membrane-spanning domain of the first repeat (IS6) region, which has been shown to be responsible for voltage-dependent inactivation. A third myotonic mutation implicates IIIS4, a region involved in sensing plasma-membrane voltage change, in the inactivation process.
Figure 4 summarizes how the EAT-2 nicotinic acetylcholine receptor, the CCA-1 T-type Ca2+ channel, and the EGL-19 L-type Ca2+ channel could cooperate to produce the upstroke of the pharyngeal muscle action potential.
|
Figure 4: The upstroke of the action potential. Schematic showing how EAT-2, CCA-1, and EGL-19 cooperate to produce the upstroke of the pharyngeal action potential. Reproduced, with permission, from Steger et al. (2005).
The timing of pharyngeal muscle relaxation is important for proper trapping of bacteria (Avery and Shtonda, 2003). A bilaterally symmetric pair of pharyngeal motor neurons, the M3s, which synapse on the corpus muscle (Albertson and Thomson, 1976), modulate the timing of relaxation. The M3s generate fast inhibitory postsynaptic potentials (IPSPs) in the contracted pharyngeal muscle. When the M3 neurons are ablated with a laser, the IPSPs disappear and the muscle contractions have a longer average duration (Avery, 1993; Raizen and Avery, 1994). Two genes were discovered to be necessary for M3→muscle transmission. eat-4 is necessary presynaptically in all glutamatergic neurons and encodes a vesicular glutamate transporter, which loads glutamate into synaptic vesicles (Bellocchio et al., 2000; Dent et al., 1997; Lee et al., 1999; Raizen and Avery, 1994). avr-15, a gene that confers sensitivity to the drug ivermectin on worms, is necessary postsynaptically for a functional M3 synapse and for the hyperpolarizing effect of glutamate on pharyngeal muscle (Dent et al., 1997). avr-15 encodes two alternatively spliced channel subunits that share ligand binding and transmembrane domains and are members of the family of glutamate-gated chloride channel subunits. An avr-15-encoded subunit forms a homomeric channel that is ivermectin-sensitive and glutamate-gated. These results indicate that: (1) an ivermectin-sensitive chloride channel mediates fast inhibitory glutamatergic neuromuscular transmission; and (2) a nematocidal property of ivermectin derives from its activity as an agonist of glutamate-gated chloride channels in essential excitable cells such as those of the pharynx. In fact, AVR-15 is one of three targets of ivermectin; the others are encoded by avr-14 and glc-1, which also encode glutamate-gated chloride channel subunits. Ivermectin acting on any of these three is sufficient to kill a worm (Dent et al., 2000). A crystal structure of ivermectin bound to GLC-1 was recently published (Hibbs and Gouaux, 2011).
AVR-15 is a member of the family of ligand-gated chloride channels, which also includes vertebrate glycine and GABA-A receptors (Arena et al., 1991; Dent et al., 1997). Although glutamate-gated chloride channels are found in many animals, vertebrates lack them, and the vertebrate ligand-gated chloride channels are far less sensitive to ivermectin than are glutamate-gated chloride channels. This explains why ivermectin is relatively harmless to mammals even though it is effective against nematodes and some other parasitic animals.
The most unusual of the pharyngeal ion channels is EXP-2. EXP-2 is a member of the Kv family of voltage-gated potassium channels. The phenotypes of loss-of-function and gain-of-function mutations (Davis et al., 1999) suggested that exp-2 encodes a subunit of the negative spike channel which mediates fast repolarization of pharyngeal muscle, first identified in the pharynx of the large parasitic nematode Ascaris by Byerly and Masuda (1979). Voltage-clamp recordings confirm this: wild-type C. elegans pharynxes have an outward current whose properties match those of Byerly and Masuda's negative spike current (Shtonda and Avery, 2005) and of EXP-2 expressed in oocytes (Fleischhauer et al., 2000)–it turns on when voltage changes from positive to negative. An exp-2 null mutant lacks this current (Davis et al., 1999; Shtonda and Avery, 2005).
exp-2 was first identified on the basis of a gain-of-function mutation that changes a cysteine at position 480 in the S6 transmembrane helix to a tyrosine (Reiner and Thomas, 1995; Thomas, 1990 ). In oocytes the mutant channel was unable to close fully, explaining the mutant phenotype (Davis et al., 1999). Modeling the S5/pore/S6 region of EXP-2 on the Kcsa structure suggested that the rotation of the S6 helix that accompanies channel closing (Perozo et al., 1999) was compatible with a cysteine at position 480, but would be blocked by a large tyrosine side chain bumping into the S5 helix. Espinosa et al. (2001) showed that indeed small amino acids at position 480 give wild-type channel function, but large ones prevent channel closing. Replacement of a conserved glycine in the S5 helix opposite C480 with bulky amino acids also produces a channel that cannot close fully.
Channels of the Kv family open when the membrane depolarizes. Typically inactivation follows opening, but it is slower. Thus the channel will conduct when membrane potential goes from negative to positive (as the channel is open but not yet inactivated for a while), but not when the membrane repolarizes, since in this direction the channel closes faster than it deinactivates. EXP-2, however, inverts this logic: it activates rather slowly, but inactivates very rapidly (Fleischhauer et al., 2000). Thus, it conducts only when membrane potential goes from positive to negative. Since the opening of the channel allows potassium to exit the cell, causing membrane potential to become more negative, which causes more EXP-2 channels to open, EXP-2 mediates a fast regenerative negative-going current spike, entirely analogous to the positive-going sodium current spike that underlies the upstroke of a typical neuronal action potential (Byerly and Masuda, 1979). EXP-2 can thus remain dormant during an action potential of unpredictable length, yet efficiently return the muscle cell to resting potential when membrane potential drops below its threshold, because of M3 firing, for instance. It is thus well adapted to end action potentials of varying duration (Shtonda and Avery, 2005). Although mammals do not have a negative spike channel, a distantly related member of the Kv family, the HERG potassium channel, has a similar inversion of gating and inactivation kinetics and plays a similar role in the heart. Pharmacological or genetic disruptions in HERG function lead to long-lasting cardiac action potentials, a dangerous condition called long QT syndrome, which can lead to fatal arrhythmia (Curran et al., 1995; Sanguinetti et al., 1995; Tseng, 2001).
During a pump, the muscles of the corpus, anterior isthmus, and TB all contract together and electrophysiological recordings show that action potentials in the corpus and TB begin simultaneously (Raizen and Avery, 1994). However, as described above, during rapid pumping action potentials are initiated by MC firing and MC does not have output onto TB muscle. Furthermore, corpus and TB contractions remain synchronized even when all pharyngeal neurons are killed (Avery and Horvitz, 1989). This suggests that, like vertebrate cardiac muscle cells, C. elegans pharyngeal muscles are electrically coupled by gap junctions. In fact, dye injected into one pharyngeal muscle cell diffuses into others in seconds (Starich et al., 1996). C. elegans, however, does not have any genes encoding connexins, the subunits of most vertebrate gap junctions. This apparent contradiction was resolved with the identification of the innexins, a family of gap junction-forming proteins found in invertebrates (Phelan et al., 1998). In mutants lacking one of these, EAT-5, corpus and TB contractions are not synchronized, and dye injected into the TB does not diffuse into the corpus (Starich et al., 1996).
Interestingly, uncoupled corpus and TB muscle may be the ancestral state. Chiang et al. (Chiang et al., 2006) found that in most free-living nematode species, corpus and TB can contract independently. MC controlled the corpus in all species, but in species in which the TB was not strongly electrically coupled to the corpus, TB contractions were controlled by motor neuron M4. In wild-type C. elegans M4 controls isthmus peristalsis (see below) but has no detectable effect on TB. However, in a mutant lacking the BK Ca2+-activated potassium channel SLO-1, M4 is able to excite TB muscle (Chiang et al., 2006). This is consistent with other studies suggesting that BK channels act presynaptically to suppress synaptic transmission (Davies et al., 2003; Pattillo et al., 2001; Roberts et al., 1990; Robitaille and Charlton, 1992; Wang et al., 2001). Apparently in wild-type C. elegans M4→TB transmission is suppressed, but the ancestral state can be recovered by increasing synaptic transmission by knocking out slo-1.
The rate of pumping determines the amount of food intake and the rate of growth. Pumping rate is determined most directly by the motor neuron MC, whose firing initiates pharyngeal muscle contraction. Mutants defective in neurotransmission from MC to pharyngeal muscle have the appearance of severely starved worms and retarded growth due to the reduced pumping rate (McKay et al., 2004; Raizen et al., 1995). This has been shown by both genetic and laser killing studies (Avery and Horvitz, 1989; Raizen et al., 1995). MC is controlled by two factors: food sensed (probably mechanically) by MC itself (Avery and Horvitz, 1989; Raizen et al., 1995) and serotonin (Avery and Thomas, 1997; Hobson et al., 2006). When food is present, mechanosensation and serotonin signal redundantly on MC in order to cause fast pumping.
Electron micrograph (EM) reconstruction shows that of the 14 pharyngeal neuron types, only I1 has chemical synapses to MC (Albertson and Thomson, 1976). I1 is also the only pharyngeal neuron that receives direct inputs from the extrapharyngeal nervous system, specifically from the collection of head ganglia called the nerve ring where most neurons and their processes are concentrated, the worm equivalent of the brain. Food and serotonin can still stimulate MC when I1 is killed with a laser (Raizen et al., 1995, David Raizen, personal communication), suggesting that in the presence of food, MC is the master neuron responsible for fast pumping. However, I1 might still mediate fine tuning of MC firing. One example is I1 regulation of basal pumping rate. In the absence of food, well-fed worms pump at a low basal rate, whose purpose David Raizen (personal communication) has speculated is to sample the environment. This basal pumping rate was significantly decreased when I1 was killed with a laser. MC ablation also decreases basal pumping. Killing both MC and I1 has the same effect as killing either alone. These data together with knowledge from EM reconstructions that I1 synapses on MC suggest that basal pumping probably results from I1 stimulating MC.
In the absence of food, MC activity is normally low. However, exogenous serotonin causes MC to fire as fast in the absence of food as in the presence of food. This is one of several examples of serotonin mimicking the effects of food on behavior (Horvitz et al., 1982). Two pharyngeal serotonin receptors have been identified; SER-1 in the pharyngeal muscle (Tsalik et al., 2003) and SER-7 in MC. MC is not stimulated by serotonin in a ser-7 knockout mutant (Hobson et al., 2006). SER-7 is thus the MC serotonin receptor.
The second feeding motion, isthmus peristalsis, is less well understood. It is a posterior-moving peristaltic contraction of the muscles of the posterior half of the isthmus, which carries food accumulated in the anterior half of the isthmus back to the TB (Avery and Horvitz, 1987). This always occurs immediately following a pump, but it doesn't happen after every pump—during rapid pumping, only about one out of four pumps is followed by an isthmus peristalsis. A posterior-moving Ca2+ wave occurs in the posterior isthmus following some but not all pumps (Shimozono et al., 2004) and is likely to correspond to isthmus peristalsis, but the calcium wave has not been directly shown to correlate with contraction. The anterior half of the isthmus, in contrast, contracts with every pump, and its contraction, although slightly delayed, overlaps that of the corpus and TB (Avery and Shtonda, 2003). Posterior isthmus peristalsis is absolutely dependent on motor neuron M4, but anterior isthmus contraction is unchanged when M4 is killed (Avery and Horvitz, 1987). Furthermore, since as described above corpus and TB contractions are synchronized by electrical coupling and since the isthmus is between them, the entire isthmus must be electrically excited at every pump, and yet only the anterior isthmus contracts at this time. The isthmus contains three pm5 muscle cells, each of which runs its entire length (Albertson and Thomson, 1976), so the differing motions of the anterior and posterior isthmus are subcellular specializations.
Isthmus peristalsis is an all-or-none event and, since it depends on M4, it is tempting to speculate that an isthmus peristalsis occurs only if M4 fires an action potential. This hypothesis receives some support from electrophysiological recordings from slo-1 mutants, in which an M4 excitatory postsynaptic potential can sometimes be detected immediately following a pump, at just the time that isthmus peristalsis might occur (Chiang et al., 2006), but there is no direct evidence correlating M4 firing with isthmus peristalsis. M4 is cholinergic as shown by its expression of unc-17, which encodes the vesicular acetylcholine transporter (Alfonso et al., 1993; Varoqui et al., 1994). The acetylcholine agonist arecoline can cause posterior isthmus muscle to contract in worms whose M4 neurons have been killed with a laser, although the contractions are uncoordinated (Bomi Song, personal communication), and the M4 EPSPs seen in slo-1 mutants depend on cholinergic transmission. This suggests that M4 acts, at least in part, by releasing acetylcholine onto pharyngeal muscle. However, M4 also expresses neuropeptides (Pocock and Hobert, 2010), and it is likely that these also contribute.
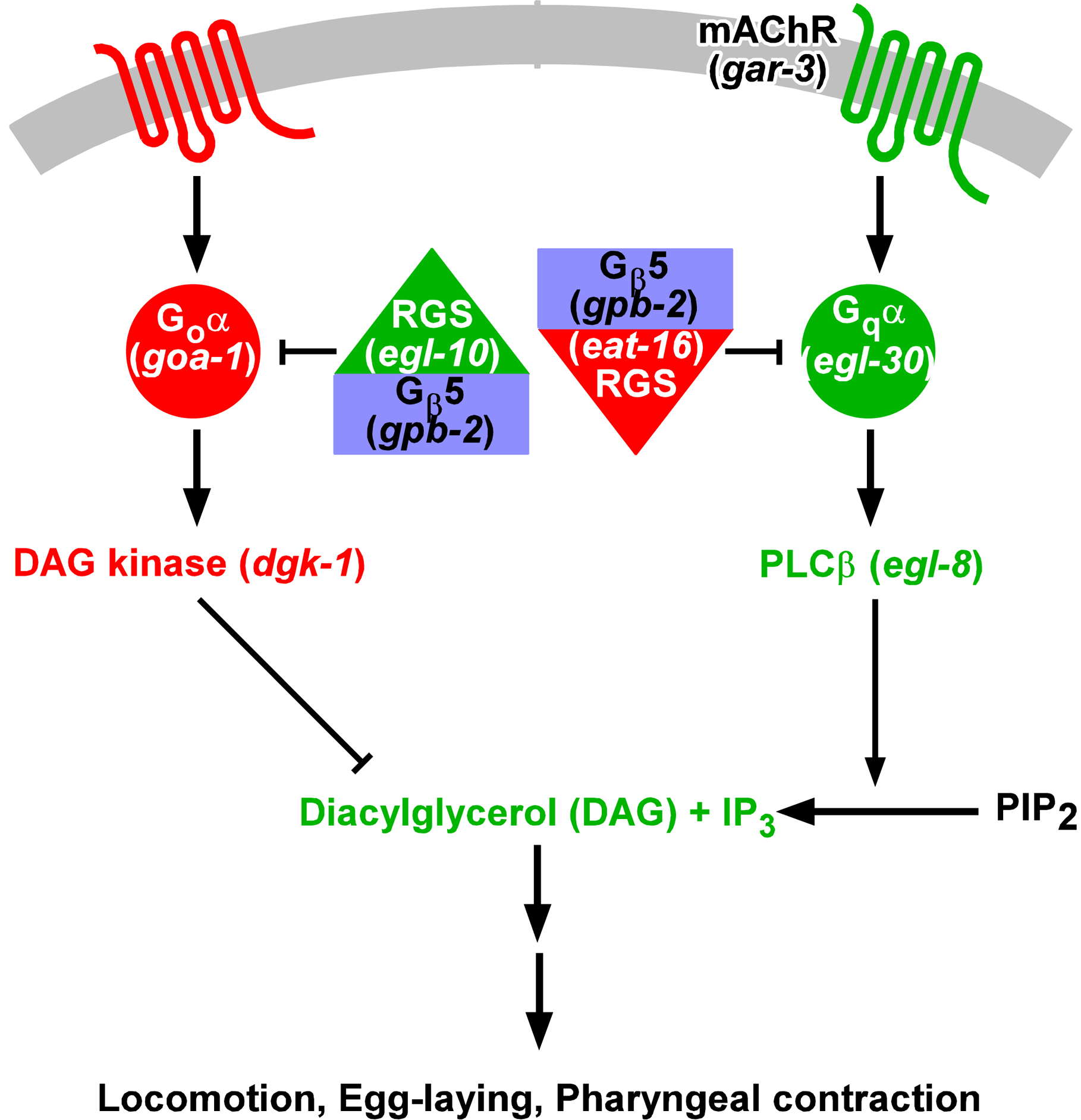
The gene eat-11, identified by a mutation that makes C. elegans hypersensitive to the muscarinic agonist arecoline (Brundage et al., 1996), was cloned by Robatzek et al. (Robatzek et al., 2001). At the same time two other labs knocked out the gene (Chase et al., 2001; van der Linden et al., 2001), which they called gpb-2. gpb-2 encodes the C. elegans ortholog of Gβ5. Vertebrate Gβ5 binds to RGS proteins that contain a G-γ-like (GGL) domain, allowing them to inhibit Gα by stimulating the intrinsic GTPase activity (Posner et al., 1999; Snow et al., 1998). C. elegans has two GGL-RGS proteins, EGL-10, which inhibits Goα, and EAT-16, which inhibits Gqα. GPB-2 is necessary for the function of both (Chase et al., 2001; Robatzek et al., 2001; van der Linden et al., 2001) (Figure 5).
|
Figure 5: Gqα/Goα mutual inhibition through Gβ5/RGS complexes. This figure shows the components of the mutually inhibitory Gqα/Goα signaling pathways that operate in many C. elegans cells. The names of the genes encoding them are shown in parentheses. Elements in green (mAChR, Gqα, EGL-10 RGS, PLCβ, DAG, and IP3) promote Gqα signaling; elements in red (Goα, EAT-16 RGS, DAG kinase, and the hypothetical serpentine receptor that activates Goα) oppose Gqα signaling. Gβ5 (in blue) is necessary for the action of both RGSs. Reproduced, with permission, from Robatzek et al. (2001).
Gqα and Goα are opposing elements of a regulatory circuit that operates in many C. elegans cells. The action of Gqα is normally restrained, directly by the GPB-2/EAT-16 RGS complex, and indirectly by Goα inhibition of its downstream effects through diacylglycerol kinase DGK-1. Steger and coworkers (Robatzek et al., 2001; Steger and Avery, 2004) showed that gpb-2 is hypersensitive to the muscarinic agonist arecoline because of unrestrained Gqα activation in pharyngeal muscle in gpb-2 mutants. There are three muscarinic receptor genes in the C. elegans genome: gar-1, gar-2, and gar-3, and Steger and Avery identified GAR-3 as the relevant muscarinic receptor by isolating a gar-3 mutation as a suppressor of the gpb-2 hypersensitivity to arecoline. They also found that atropine, a specific muscarinic antagonist, protects gpb-2(ad541) worms against arecoline, confirming that hypersensitivity requires muscarinic receptor activation. Furthermore, overexpression of GAR-3 makes worms hypersensitive to arecoline. In support of the GAR-3 function in pharyngeal muscle as a Gq coupled receptor, a gar-3 reporter fusion is expressed in pharyngeal muscle, and GAR-3 expressed in heterologous systems activates phospholipase C (PLC) and protein kinase C (PKC) (Lee et al., 2001; Min et al., 2000). gar-1 and gar-2 mutations have no effect on arecoline hypersensitivity.
The immediate effect of the GAR-3 pathway is to increase muscle contraction, by increasing both the duration of the muscle action potential and the strength of contraction (Steger and Avery, 2004). In gar-3 mutants the TB action potential is briefer than in wild type. In gpb-2(ad541) worms, arecoline causes the pharynx to contract and stay contracted for as long as a minute, and a gar-3 mutation prevents this hypercontraction. However, EPGs show that action potentials continue during the hypercontraction. That the pharynx remains contracted between action potentials under these conditions shows that GAR-3 also increases contraction in a membrane potential-independent way.
There are no data that suggest a gar-1 function in the pharynx. However, gar-2 is likely to function in neurons for the following two reasons: (1) A gar-2::GFP fusion is expressed exclusively in the nervous system (pharyngeal and extrapharyngeal); and (2) gar-2 mutants are hypersensitive to the acetylcholinesterase inhibitor aldicarb, an extensively characterized phenotype that results from increased synaptic release of acetylcholine (Rand and Nonet, 1997). This suggests that GAR-2 inhibits acetylcholine release. Most neurons contain receptors for their own neurotransmitters that mediate feedback inhibition—this may be one function of GAR-2.
Hunger is the internal state that results from starvation and that motivates the behavioral response. Hungry animals seek food, increase exploratory behavior, increase alertness and feed when food is available. What mediates the changes in behavior? You et al. found that the muscarinic receptor → MAPK (Mitogen-Activated Protein Kinase) pathway is part of hunger signaling in the pharynx (You et al., 2006): the activation of this pathway by starvation can be detected by phospho-specific antibody to MAPK. Muscarinic signaling during starvation causes pharyngeal muscle to undergo specialized changes, which probably prepare it to encounter food later. Moreover, overactivation of this pathway in the sensitized mutant gpb-2 causes lethality due to a pharyngeal muscle change that directly interferes with feeding motions, suggesting that fine-tuned regulation of this pathway is essential for worm survival during starvation.
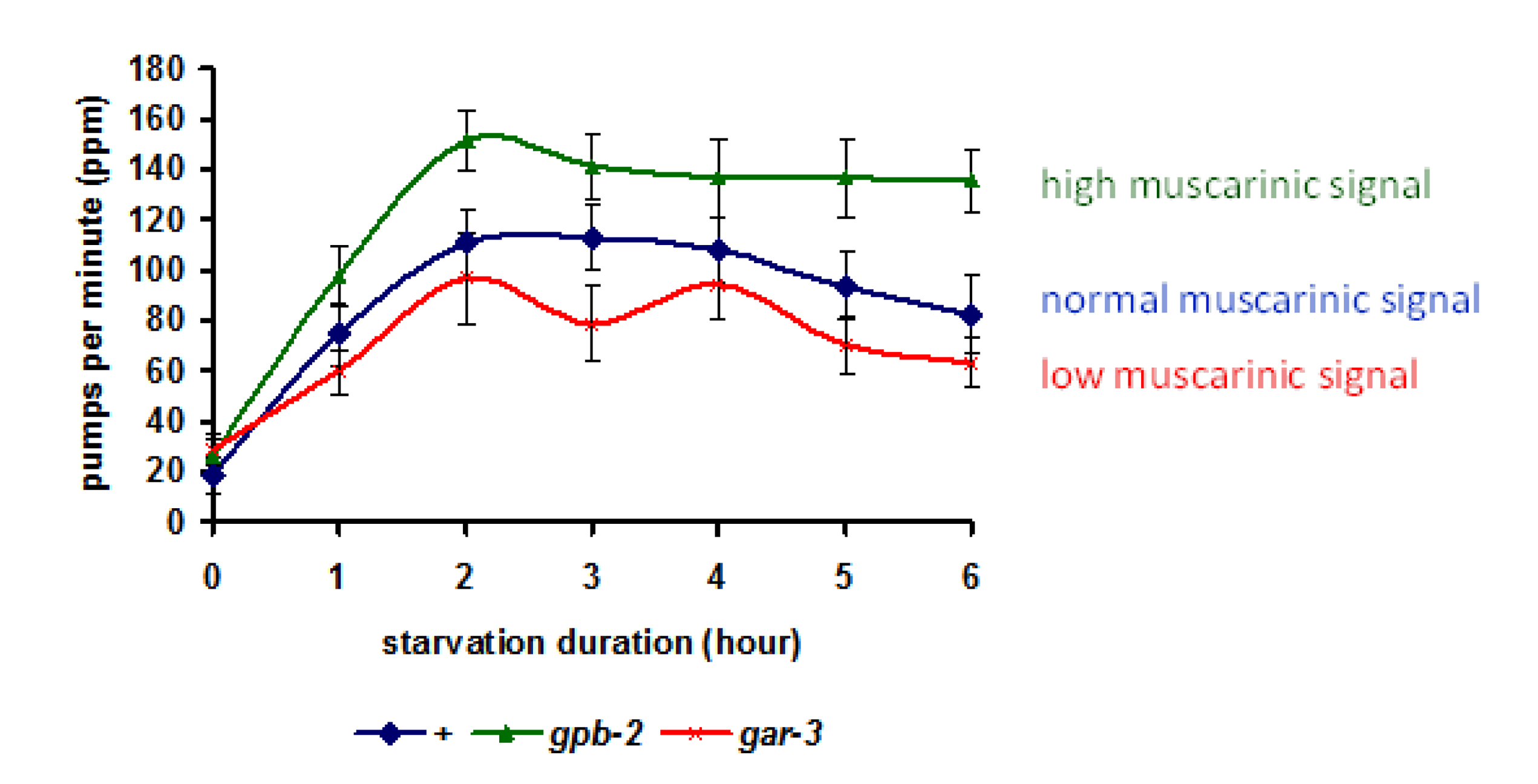
To understand the purpose of the muscarinic signaling to MAPK pathway during starvation, You et al. measured pumping rate (Figure 6). Wild type worms initially pump slowly when they are taken off food, but they increase pumping rate gradually in the first 2 hours of starvation. When the pharyngeal muscle GAR-3→MPK-1 pathway is blocked with a gar-3 mutation, the increase in pumping rate is reduced. Conversely, in gpb-2 mutants, in which the pathway is hyperactive, pumping rate increases more on starvation. These data suggest that activation of the muscarinic receptor during starvation contributes to the increase in starvation-induced pharyngeal activity.
|
Figure 6: Starvation induces pumping in a muscarinic signaling-dependent manner. Pumping rates increased for first 2 hr in the absence of food. Two mutants defective in muscarinic signaling showed altered pumping rates compared to wild-type. gpb-2 mutants, which have upregulated muscarinic signaling, showed a higher pumping rate, and gar-3 mutants, which have downregulated muscarinic signaling,showed a lower pumping rate than wild-type. Reproduced, with permission, from You et al. (2006).
By what molecular mechanisms does the activation of muscarinic receptor → MAPK pathway allow worms to cope with starvation? Kang et al. (Kang et al., 2007) found that the pathway activates autophagy, helping worms to survive starvation, and that excessive autophagy during starvation in gpb-2 mutants kills the animals, contributing to their hypersensitivity to starvation.
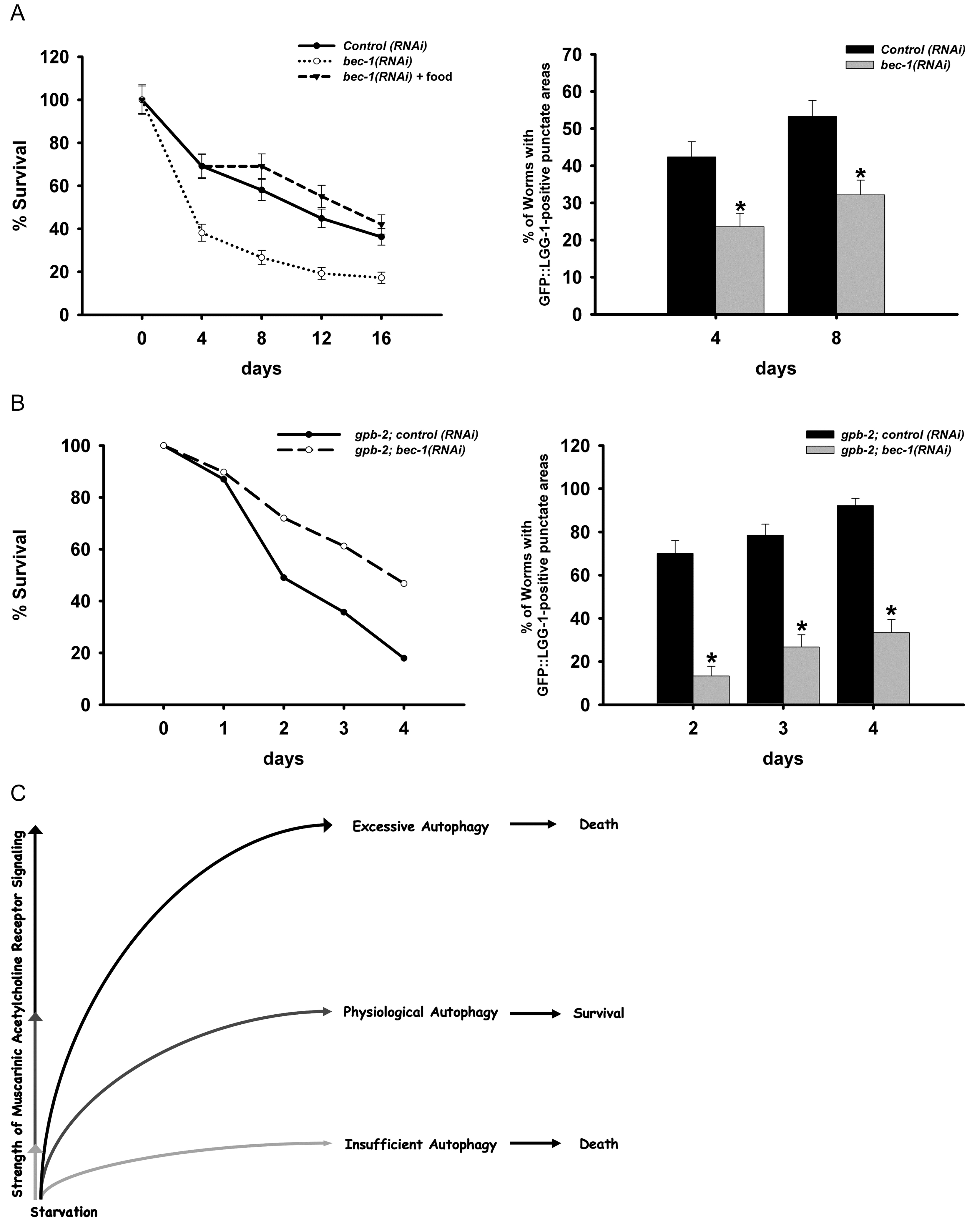
Autophagy is a major pathway used to degrade long-lived proteins and organelles. It is thought to promote both cell and organism survival by providing fundamental building blocks to maintain energy homeostasis during starvation. Under different conditions, however, autophagy may instead act to promote cell death through an autophagic cell death pathway distinct from apoptosis. Kang et al. (Kang et al., 2007) found that autophagy can also promote survival or cause death of the whole organism. Physiological levels of autophagy act to promote survival in C. elegans during starvation, whereas insufficient or excessive levels of autophagy contribute to death (Figure 7). When autophagy is inhibited during starvation, it decreases survival of wild-type worms (Figure 7). In contrast, in gpb-2 mutants, in which muscarinic signaling cannot be down-regulated, starvation induces excessive autophagy in pharyngeal muscles, which in turn, causes damage that may contribute to death (Figure 7B).
|
Figure 7: An optimal level of autophagy for starvation survival. A: Autophagy promotes starvation survival. When autophagy is inhibited in starved wild-type worms by bec-1 RNAi, they are unable to survive as long. B: Excess autophagy causes premature death. gpb-2 worms have excess autophagy due to unrestrained muscarinic signaling and die rapidly during starvation. In this background inhibition of autophagy promotes survival. C: Summary of effect of level of autophagy on starvation survival. Adapted, with permission, from Kang et al. (2007).
Shtonda and Avery (Shtonda and Avery, 2006) showed that worms are able to distinguish food based on quality (operationally defined as ability to support growth) and to seek out higher quality food.
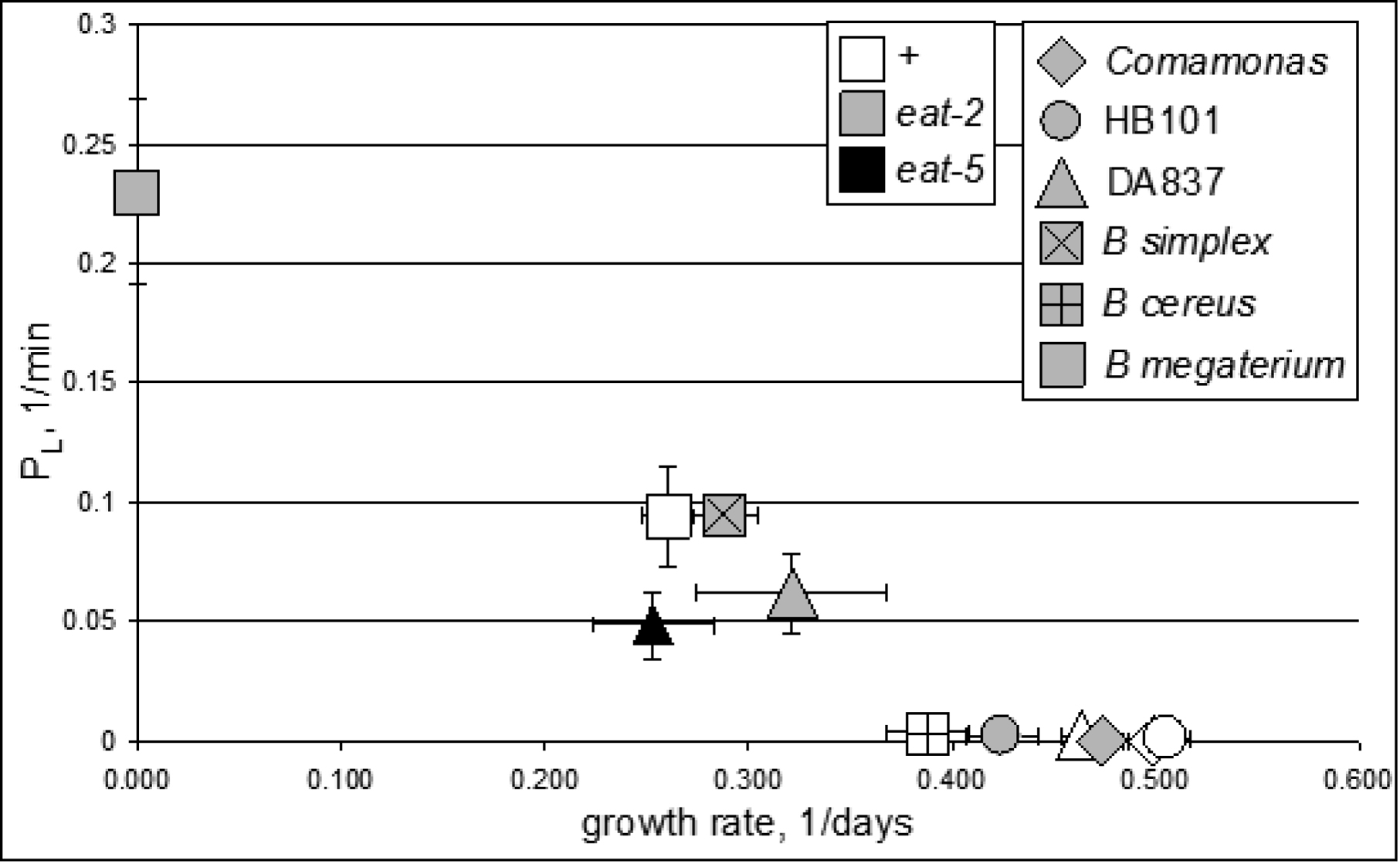
Hydrodynamic simulation (http://elegans.som.vcu.edu/~leon/pharynx_sim/; Avery and Shtonda, 2003) to understand how bacteria are transported within the pharynx led to a prediction: large bacteria would be poor food. Shtonda (Avery and Shtonda, 2003; Shtonda and Avery, 2006) isolated bacteria from soil samples and tested them for their ability to support C. elegans growth. He defined food quality as the ability of a food to support C. elegans growth, as measured by growth rate: the inverse of the time for a worm to grow from hatching to adulthood. (Growth rates range from approximately 0.48/day on the best food to 0.) Figure 8 shows the almost perfect inverse correlation between bacterial size and food quality: big bacteria are poor food. There is one conspicuous exception: E. coli strain DA837. It is the same size as E. coli HB101 but a far worse food source because DA837 bacteria clump, making the effective size of the bacteria bigger than an individual bacterium.
|
Figure 8: Large bacteria are poor food. The bar graph shows growth rates of wild-type worms and three different feeding-defective mutants on 16 different bacterial strains. The bacteria are listed in order of decreasing eat-2 growth rate. Above the bar graph are photographs of each of the bacterial strains as they appear when isolated from lawns on nematode growth medium. All pictures are on the same scale; a scale bar is shown to the left. In addition, 0.8 μm blue-dyed latex beads were mixed in with the bacteria as an internal size standard. These are identified by black dots. A strong although not perfect inverse correlation between growth rate and bacterial size is obvious. Reproduced, with permission, from Avery and Shtonda (2003).
Shtonda and Avery (Shtonda and Avery, 2006) characterized two behaviors that allow worms to find high quality food: (1) leaving, in which a worm leaves a poor food in search of better, and (2) preference, in which worms given a choice between two foods will choose the higher quality one. Worms that have found food are reluctant to leave, but if the food is of poor quality, they will do so. Figure 9 shows that naive worms (hatched in the absence of food and starved until the assay) will leave food that can't support growth at a rate of 0.4/day or better. Time courses show that leaving probability PL reaches its steady-state value within ten minutes, suggesting that assessment of food quality happens quickly. PL depends on the worms as well as the food: eat-2 and eat-5 mutants show a substantial probability of leaving DA837, while wild-type worms will not. Since the direct effects of mutations in eat-2 and eat-5 are specific to the pharynx, this suggests that assessment of food quality depends on something that occurs in or past the pharynx.
|
Figure 9: Leaving probability depends on food quality. This graph summarizes 11 leaving probability measurements. In each experiment naive hatchlings starved for about a day were placed 2 mm from an elliptical 1 × 2 mm colony of bacteria and videotaped. PL is determined for each minute as the ratio of the number of worms leaving the colony to the total number of worms in the colony. The average over the time from 30 to 90 min is plotted here. Each point is color-coded for worm strain and shaped-coded for bacterial strain as indicated in the legends. Reproduced, with permission, from Shtonda and Avery (2006).
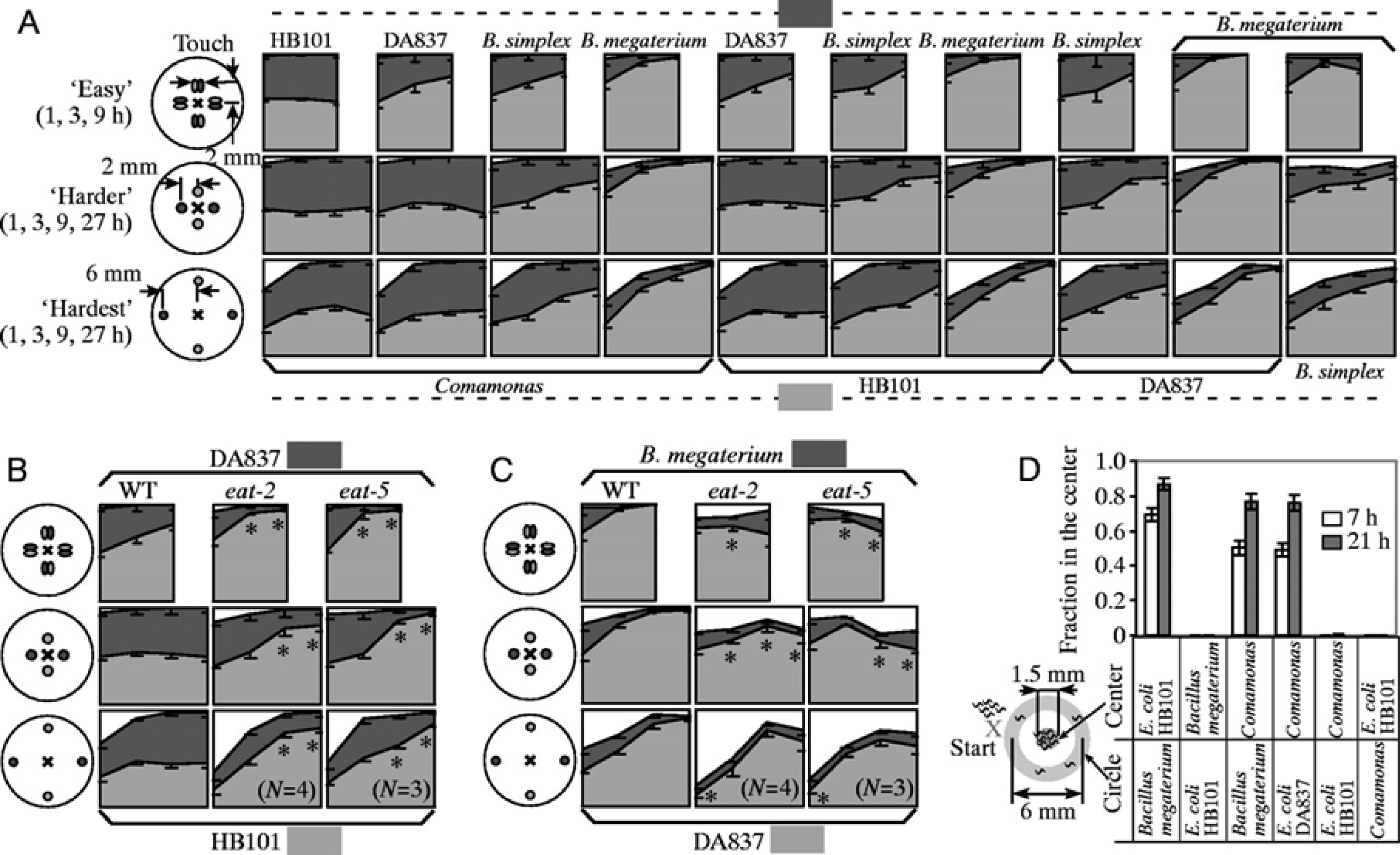
To test whether worms would seek out better food (showing preference), Shtonda (Shtonda and Avery, 2006) placed several hundred naïve L1s on plates containing spots of two different types of bacteria. The distribution of worms at later times is shown in Figure 10. Without exception, more worms are found in the better food. The strength of the preference usually correlates with the disparity in food quality. Significantly, preference is usually higher for eat-2 and eat-5 mutant worms than for wild-type and is never lower.
|
Figure 10: Worms choose higher quality food. These graphs show the result of experiments in which worms were given the choice between two different foods. In A-C, small spots of bacteria of two different types were placed on a plate in the arrangements shown, then naïve worms were placed in the center of the plate. After 1, 3, 9, and 27 hours, each worm was scored to see whether it was inside a bacterial spot, and if so, which type of bacteria. In each plot light gray is the percentage of worms in the higher quality food, dark gray those in lower quality food, and white those outside the food. A. Wild-type worms choosing between pairs selected from four different bacterial strains. B. Wild-type, eat-2, and eat-5 worms choosing between E. coli strains HB101 and DA837. C. Wild-type, eat-2 and eat-5 choosing between E. coli DA837 and Bacillus megaterium. D. A different task, in which worms are required to leave one food source to find another. One strain of bacteria was placed in the center of a plate, while a second strain was placed in e circle completely surrounding the center spot. Worms were placed outside the circle, then the proportion in the center spot were scored after 7 or 21 hours. When the circle was made of low-quality food, worms were able to cross it to find high-quality food in the center. Reproduced, with permission, from Shtonda and Avery (2006).
A simple explanation for preference would be that it is just a consequence of differences in leaving probability. If worms leave poor food frequently and good food rarely, they will, on average, spend more time in the good food than the poor. But a quantitative argument shows that differences in naïve leaving probability cannot be the whole story. Preferences can be very strong: ratios near 100 are routinely observed. For instance, eat-2 mutants show a 200-fold preference for DA837 over B. megaterium. The ratio of naïve PL's, however, is only 0.23/0.062 = 3.7 (Figure 9). The accumulation in the better of two foods cannot be explained solely by the reactions of the worms to the individual foods considered separately. Somehow a comparison is being made.
Worms modify their behavior depending on their previous experience of food quality (Avery and Shtonda, 2003; Shtonda and Avery, 2006). Worms grown for 3 hrs on high-quality food then presented with medium-quality food were dissatisfied—they left the mediocre food at high frequency. But when worms were presented with the same mediocre food after conditioning on low-quality food they were satisfied—they left the mediocre food at low frequency (Figure 11A). Likewise, in a biased preference assay in which worms are required to cross a ring of medium quality food to find good food, worms conditioned on high-quality food performed better (Figure 11B). Furthermore, Shtonda found that the extrapharyngeal interneuron AIY is critical for this behavior. When AIY was inactivated either by mutation or laser-killing, the worms were strongly biased towards dwelling (the food-exploiting locomotory behavior (Fujiwara et al., 2002)) and away from roaming (food-seeking), as if they perceived the food to be of higher quality than it was (Shtonda and Avery, 2006).
|
Figure 11: Food-seeking depends on experience. A. Naïve L1s were conditioned for 3 hours on the indicated bacteria then their probability of leaving a small spot of medium quality food E. coli DA837 was measured as in Figure 9. “Empty plate” means the worms were left on a plate without food for 3 hours, instead of being conditioned on bacteria. Worms that have experienced high quality food leave medium quality food at higher frequency. B. After conditioning for 2 hours, worms were tested for their ability to cross a ring of medium quality food to find a spot of high-quality food in the center. Worms conditioned on high-quality food performed this task better. Adapted, with permission, from Shtonda and Avery (2006).
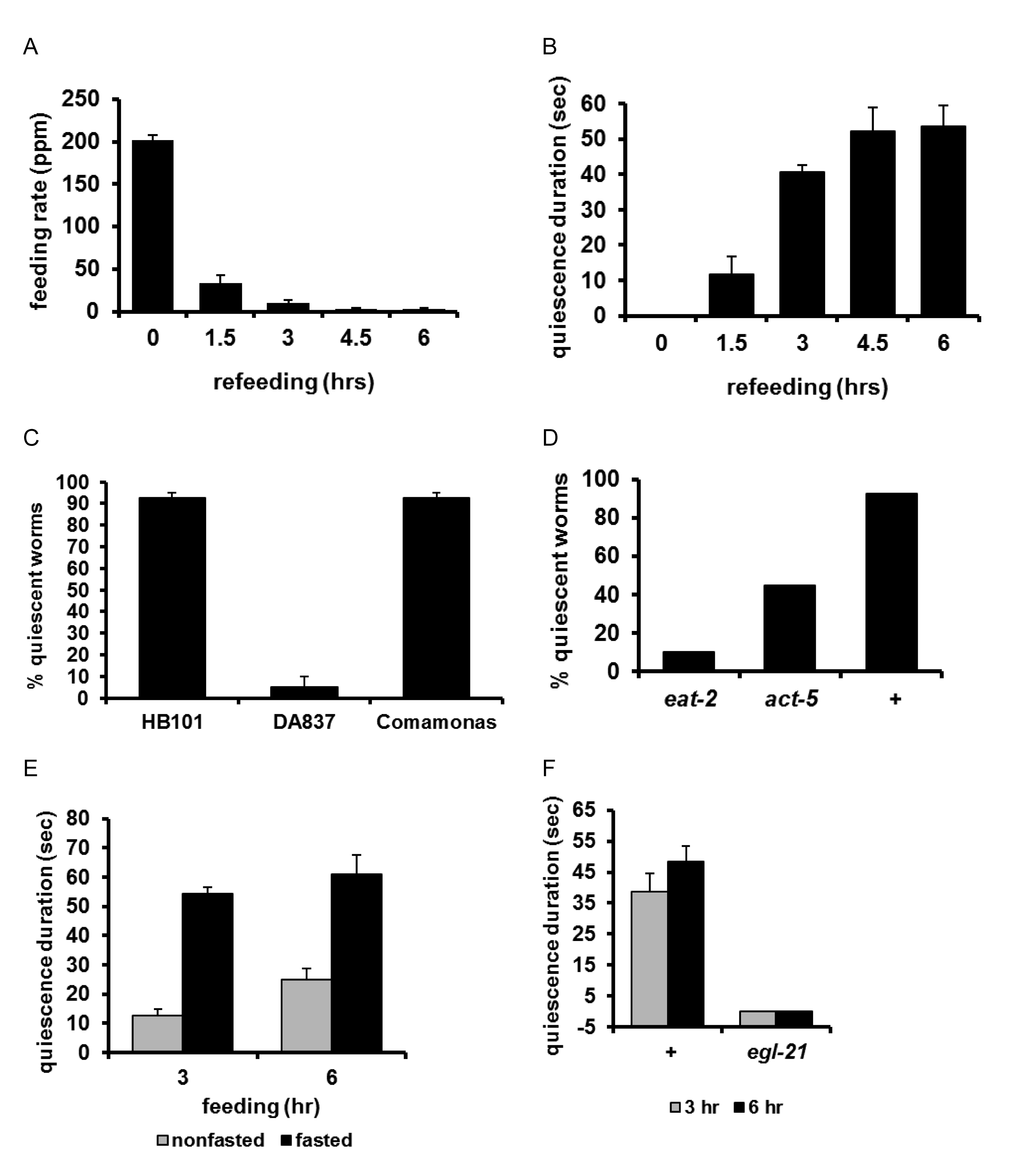
Shtonda's discovery that worms respond to food quality led to the discovery of another feeding behavior called satiety quiescence. Satiated animals of various species stop eating, decrease exploratory behavior and often fall asleep, a pattern called the ‘behavioral sequence of satiety’ (Antin et al., 1975). No such behavior had been reported in C. elegans, however. Most worm labs fed worms E. coli OP50 (of which DA837 is a derivative), which turns out to be relatively poor quality food. You et al. (You et al., 2008) found that under certain conditions, one of the more important being high quality food, worms display the behavioral sequence of satiety. When satiated, they stop eating (measured by pumping rate), stop moving, and often become quiescent (movie and Figure 12A,B). You et al. called this behavior ‘satiety quiescence’ because: (1) The quiescence is dependent on food quality–worms become quiescent on good food but not on poor food (Figure 12C); (2) decrease in food intake (in feeding mutants such as eat-2) or decrease in food absorption in the intestine (in absorption mutants such as act-5) reduces quiescence (Figure 12D); and (3) The behavior is dependent on the animal's past experience of starvation–worms that have experienced starvation show enhanced satiety quiescence compared to worms that have not (Figure 12E). Satiety quiescence is regulated by peptide signals since egl-21 mutants, which don't produce most peptide signals (Husson et al., 2007), are completely defective in satiety quiescence (Figure 12F).
|
Figure 12: Satiety quiescence. A, B. Worms were starved for 12 hours, then placed on high-quality food (E. coli HB101). With time they stopped eating (A) and became quiescent (B). C. Quiescence depends on food quality. Most worms fed good foods E. coli HB101 or Comamonas become quiescent; few worms fed the relatively poor food E. coli DA837 do. D. Quiescence depends on food intake. A mutation in eat-2, which slows the rate of feeding, or act-5, which decreases nutrient absorption in the intestine, reduces quiescence. E. Fasting prior to full feeding increases quiescence. Fasted worms were left for 12 hours without food before feeding on high-quality food for the indicated times. F. Quiescence depends on peptide signals. A mutation in egl-21 that prevents the processing of most peptide transmitters completely blocks quiescence. (Note that the y-axis has been displaced to allow bars of 0 height to be visible). An egl-3 mutation that also blocks almost all peptidergic transmission (Husson et al., 2006) has the same effect (not shown). Reproduced, with permission, from You et al. (2008).
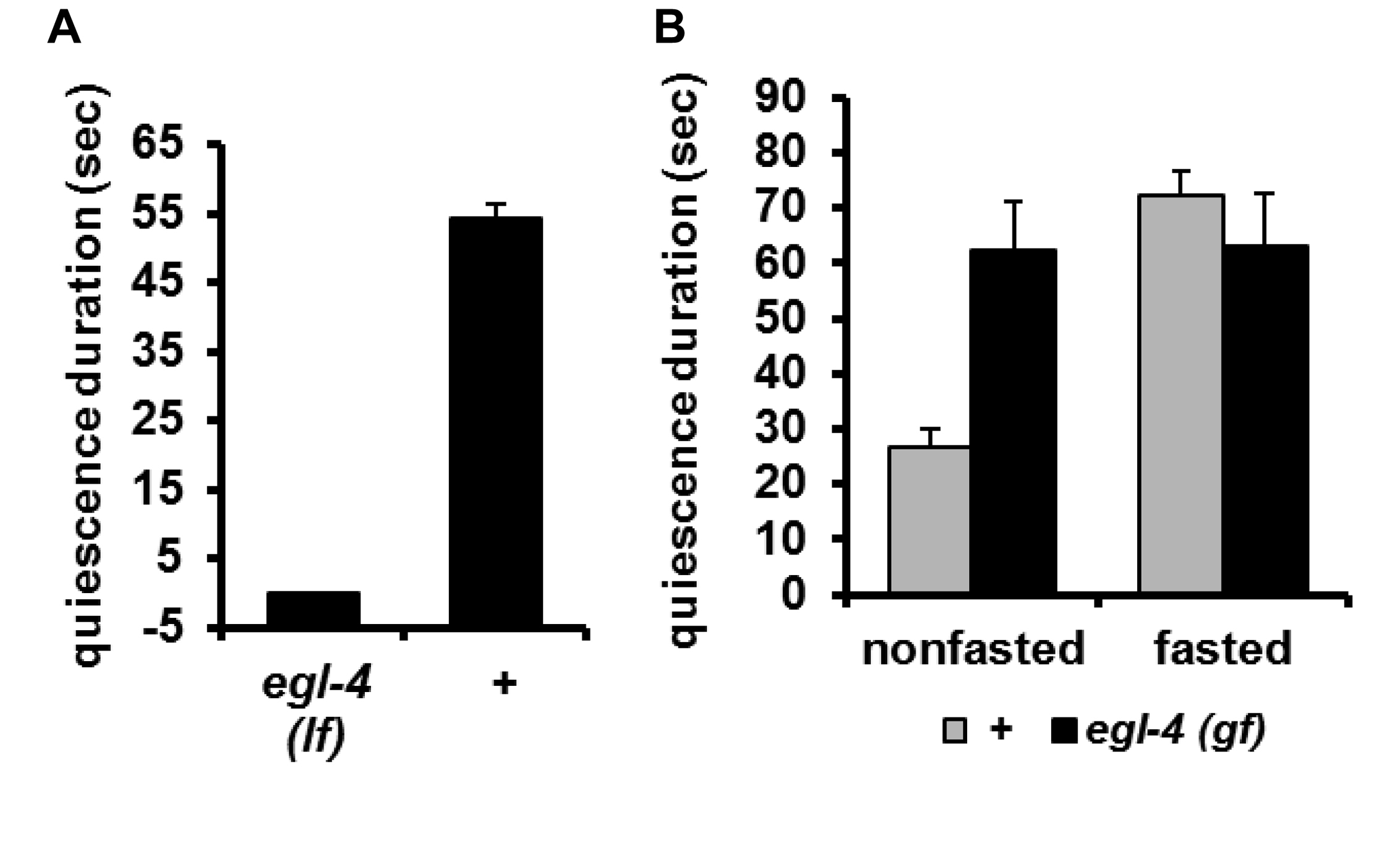
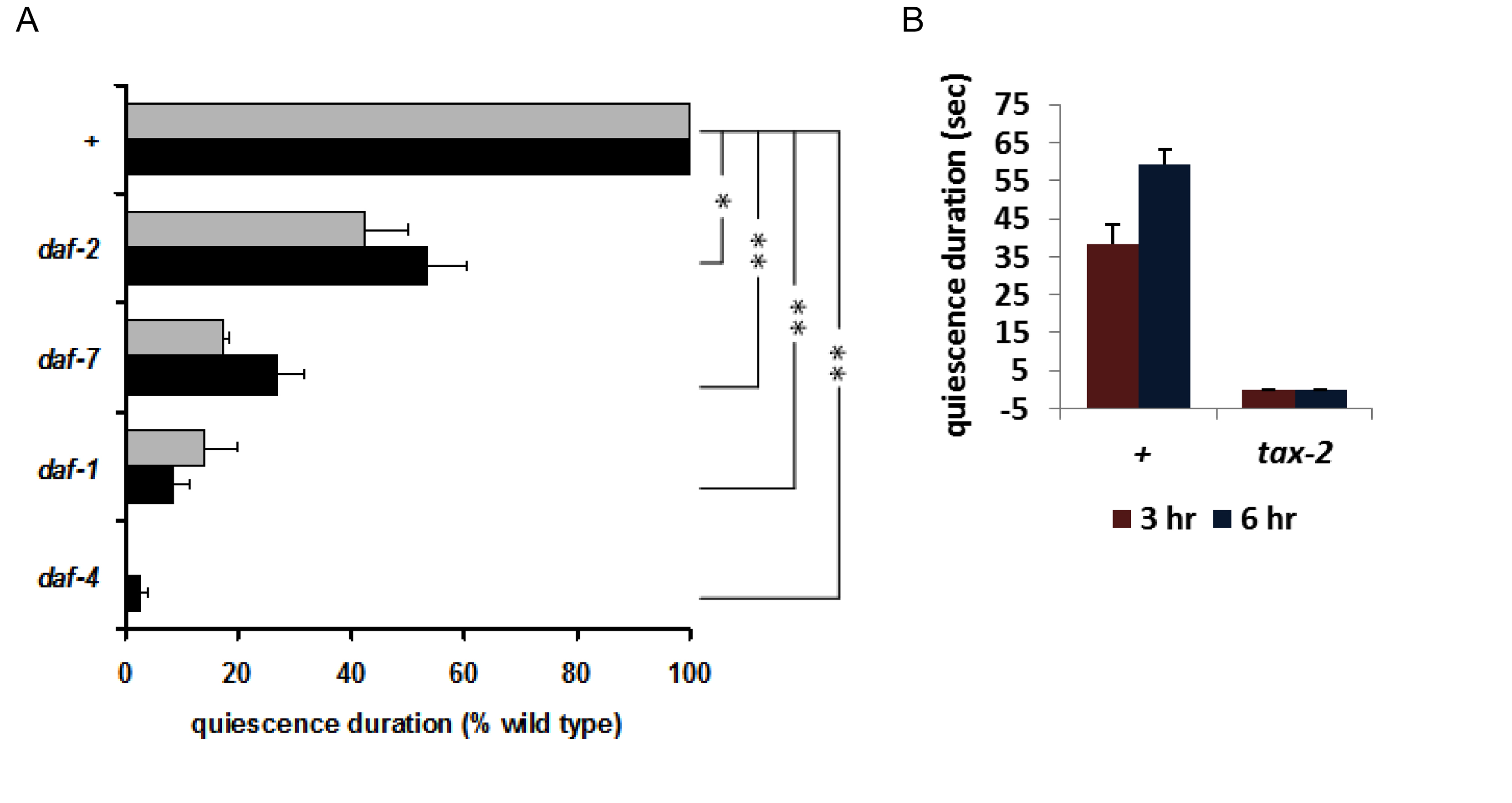
Intriguingly, this sleep-like behavior was not completely unprecedented. In a screen for feeding-defective mutants Avery (Avery, 1993) isolated mutants that had a starved appearance, although there was no obvious defect in feeding motions or the feeding rate. However, whenever he examined them after a long period in which they had not been disturbed, he found them to be completely quiescent, neither eating nor moving. He speculated that they appeared starved because they were quiescent most of the time and as a result didn't eat as much as wild type. This mutation was eventually mapped to pkg-1 (formerly egl-4), which encodes cyclic GMP-dependent Protein Kinase (PKG) (Fujiwara et al., 2002; Raizen et al., 2006). You et al. (You et al., 2008) found that pkg-1 loss of function mutants show no quiescence, whereas the gain of function mutation isolated by Avery shows excessive quiescence (Figure 13). This finding suggested a role for cGMP signaling in satiety quiescence, confirmed by the observation that the membrane guanylate cyclase DAF-11 and the cGMP-gated cation channel are necessary for satiety quiescence (Figure 14B). Consistent with the evidence for peptide signaling, insulin and TGFβ signals are also necessary for worms to show satiety quiescence (Figure 14A). In C. elegans, insulin, TGFβ and cGMP pathways are involved in sensing a favorable environment, and in making the decision to keep growing and reproducing instead of becoming a dauer, a non-reproductive form specialized for long-term survival (Riddle et al., 1981). In other words, these signals are used to ensure that worms will be in nutritionally favorable conditions. You et al.'s findings imply that these same signals of good conditions control satiety and quiescence in adults.
|
Figure 13: Satiety quiescence is dependent on pkg-1. A. A loss-of-function mutant of pkg-1, which encodes a cGMP dependent protein kinase, shows no satiety quiescence. B. A pkg-1 gain-of-function mutant shows satiety quiescence even without prefasting. Reproduced, with permission, from You et al. (2008).
Expressing pkg-1 under the control of a tax-4 promoter rescued the quiescence defect of pkg-1 mutants. tax-4 encodes a cGMP-gated channel expressed in a dozen neuron types that sense the worm's environment and internal state (Komatsu et al., 1996). These neurons have critical roles in controlling body fat, determining body size and changing behaviors depending on environmental factors such as oxygen concentration (Coates and de Bono, 2002; Fujiwara et al., 2002; Mak et al., 2006), temperature (Ramot et al., 2008), and light (Ward et al., 2008).
TAX-4 and TAX-2 are α and β subunits of a cGMP-gated channel (Komatsu et al., 1996) and their neuronal expression overlaps. When You et al. tested tax-2 mutants, they showed no quiescence (Figure 14B). (They were unable to test tax-4 mutants because, unlike tax-2, they don't stay on food.) These results suggest that the TAX-2/4 expressing neurons and their neuronal activities mediated by the TAX-2/4 cGMP-gated channel are necessary for quiescence after fasting and refeeding.
|
Figure 14: Insulin, TGFβ, and cGMP signaling is necessary for normal satiety quiescence. A. Mutations in genes encoding the insulin receptor DAF-2, the TGFβ DAF-7, and DAF-7 receptor subunits DAF-1 and DAF-4 reduce quiescence. B. TAX-2, a subunit of the cGMP-gated cation channel, is necessary for satiety quiescence. Reproduced, with permission, from You et al. (2008).
Of the TAX-2/4 expressing neurons only ASIs express DAF-7 (Ren et al., 1996), and the quiescence defect of daf-7 mutants is fully rescued by expressing daf-7 in ASI (You et al., 2008). Unfavorable conditions such as shortage of food, high temperature and high concentration of dauer pheromone induce dauer formation in worms. Worms become dauers even under favorable conditions if ASI is killed (Bargmann and Horvitz, 1991). Recent findings suggest that ASI is necessary for life span extension by diet restriction (Bishop and Guarente, 2007). These results suggest a role of the ASI neurons in regulating or sensing food intake. That quiescence after fasting and refeeding is mediated by daf-7 in ASI neurons supports this role of ASI.
Interestingly, most mutants defective in satiety quiescence have darker intestines than wild type. A dark intestine usually correlates with more fat storage (McKay et al., 2003). In fact, some of these mutants, including daf-2 and daf-7, store more fat than wild type (Kimura et al., 1997). Quiescence resulting from satiety suggests that worms don't feed constantly, but rather regulate their feeding depending on nutritional status and environment. Quiescence-defective mutants that have reduced “good conditions” signaling could be defective in regulating feeding, which might cause more feeding and fat accumulation. Recent studies suggest that natriuretic peptide receptors, homologs of the guanylate cyclase DAF-11, have a role in storing and degrading fat through PKG in adipose tissue (Sengenes et al., 2000) and that TGF-β signaling in neurons regulates adiposity during exercise (Ishikawa et al., 2006). These results suggest the possibility of conserved linkage of these signaling pathways to regulation of feeding and metabolism in worms and mammals.
An animal must regulate its feeding based on internal status and external environment. Worms alter their feeding motions depending on their nutritional status: They pump more if they are hungry, and they don't pump at all if they are satiated. But feeding is not an isolated behavior. Hunger not only makes worms pump more but move more, too. They are willing to leave food they have to seek better food if past experience suggests better food might be found. The opposite is also true: satisfied worms never leave their food, often stop pumping when they are satiated and become quiescent. Understanding feeding and its regulation may help us to understand how animals think, respond and behave so as to survive in an unpredictable environment.
Albertson, D. G., and Thomson, J. N. (1976). The pharynx of Caenorhabditis elegans. Philos. Trans. R. Soc. Lond. B Biol. Sci. 275, 299-325. Abstract Article
Alfonso, A., Grundahl, K., Duerr, J. S., Han, H. P., and Rand, J. B. (1993). The Caenorhabditis elegans unc-17 gene: a putative vesicular acetylcholine transporter. Science 261, 617-619. Abstract Article
Antin, J., Gibbs, J., Holt, J., Young, R. C., and Smith, G. P. (1975). Cholecystokinin elicits the complete behavioral sequence of satiety in rats. J. Comp. Physiol. Psychol. 89, 784-790. Abstract Article
Arena, J. P., Liu, K. K., Paress, P. S., and Cully, D. F. (1991). Avermectin-sensitive chloride currents induced by Caenorhabditis elegans RNA in Xenopus oocytes. Mol. Pharmacol. 40, 368-374. Abstract
Avery, L. (1993). The genetics of feeding in Caenorhabditis elegans. Genetics 133, 897-917. Abstract
Avery, L. (1993). Motor neuron M3 controls pharyngeal muscle relaxation timing in Caenorhabditis elegans. J. Exp. Biol. 175, 283-297. Abstract
Avery, L., and Horvitz, H. R. (1987). A cell that dies during wild-type C. elegans development can function as a neuron in a ced-3 mutant. Cell 51, 1071-1078. Abstract Article
Avery, L., and Horvitz, H. R. (1989). Pharyngeal pumping continues after laser killing of the pharyngeal nervous system of C. elegans. Neuron 3, 473-485. Abstract Article
Avery, L., and Shtonda, B. B. (2003). Food transport in the C. elegans pharynx. J. Exp. Biol. 206, 2441-2457. Abstract Article
Avery, L., and Thomas, J. H. (1997). Feeding and defecation. In C. elegans II, D. L. Riddle, T. Blumenthal, B. J. Meyer, and J. R. Preiss, eds. (New York, Cold Spring Harbor Press), pp. 679-716. Abstract
Bargmann, C. I., and Horvitz, H. R. (1991). Control of larval development by chemosensory neurons in Caenorhabditis elegans. Science 251, 1243-1246. Abstract Article
Bellocchio, E. E., Reimer, R. J., Fremeau, R. T., Jr., and Edwards, R. H. (2000). Uptake of glutamate into synaptic vesicles by an inorganic phosphate transporter. Science 289, 957-960. Abstract Article
Bishop, N. A., and Guarente, L. (2007). Two neurons mediate diet-restriction-induced longevity in C. elegans. Nature 447, 545-549. Abstract Article
Brundage, L., Avery, L., Katz, A., Kim, U. J., Mendel, J. E., Sternberg, P. W., and Simon, M. I. (1996). Mutations in a C. elegans Gqalpha gene disrupt movement, egg laying, and viability. Neuron 16, 999-1009. Abstract Article
Byerly, L., and Masuda, M. O. (1979). Voltage-clamp analysis of the potassium current that produces a negative-going action potential in Ascaris muscle. J. Physiol. 288, 263-284. Abstract
Chase, D. L., Patikoglou, G. A., and Koelle, M. R. (2001). Two RGS proteins that inhibit Galpha(o) and Galpha(q) signaling in C. elegans neurons require a Gbeta(5)-like subunit for function. Curr. Biol. 11, 222-231. Abstract
Chiang, J. T., Steciuk, M., Shtonda, B., and Avery, L. (2006). Evolution of pharyngeal behaviors and neuronal functions in free-living soil nematodes. J. Exp. Biol. 209, 1859-1873. Abstract Article
Coates, J. C., and de Bono, M. (2002). Antagonistic pathways in neurons exposed to body fluid regulate social feeding in Caenorhabditis elegans. Nature 419, 925-929. Abstract Article
Cook, A., Franks, C. J., and Holden-Dye, L. (2006). Electrophysiological recordings from the pharynx. WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.110.1. Abstract
Curran, M. E., Splawski, I., Timothy, K. W., Vincent, G. M., Green, E. D., and Keating, M. T. (1995). A molecular basis for cardiac arrhythmia: HERG mutations cause long QT syndrome. Cell 80, 795-803. Abstract Article
Davies, A. G., Pierce-Shimomura, J. T., Kim, H., VanHoven, M. K., Thiele, T. R., Bonci, A., Bargmann, C. I., and McIntire, S. L. (2003). A central role of the BK potassium channel in behavioral responses to ethanol in C. elegans. Cell 115, 655-666. Abstract Article
Davis, M. W., Fleischhauer, R., Dent, J. A., Joho, R. H., and Avery, L. (1999). A mutation in the C. elegans EXP-2 potassium channel that alters feeding behavior. Science 286, 2501-2504. Abstract Article
Dent, J. A., Davis, M. W., and Avery, L. (1997). avr-15 encodes a chloride channel subunit that mediates inhibitory glutamatergic neurotransmission and ivermectin sensitivity in Caenorhabditis elegans. EMBO J. 16, 5867-5879. Abstract Article
Dent, J. A., Smith, M. M., Vassilatis, D. K., and Avery, L. (2000). The genetics of ivermectin resistance in Caenorhabditis elegans. Proc. Natl. Acad. Sci. U S A 97, 2674-2679. Abstract Article
Doncaster, C. (1962). Nematode feeding mechanisms. I. Observations on Rhabditis and Pelodera. Nematologica 8, 313-320. Article
Espinosa, F., Fleischhauer, R., McMahon, A., and Joho, R. H. (2001). Dynamic interaction of S5 and S6 during voltage-controlled gating in a potassium channel. J. Gen. Physiol. 118, 157-170. Abstract Article
Fang-Yen, C., Avery, L., and Samuel, A. D. (2009). Two size-selective mechanisms specifically trap bacteria-sized food particles in Caenorhabditis elegans. Proc. Natl. Acad. Sci. U S A 106, 20093-20096. Abstract Article
Fleischhauer, R., Davis, M. W., Dzhura, I., Neely, A., Avery, L., and Joho, R. H. (2000). Ultrafast inactivation causes inward rectification in a voltage-gated K(+) channel from Caenorhabditis elegans. J. Neurosci. 20, 511-520. Abstract
Franks, C. J., Holden-Dye, L., Bull, K., Luedtke, S., and Walker, R. J. (2006). Anatomy, physiology and pharmacology of Caenorhabditis elegans pharynx: a model to define gene function in a simple neural system. Invert. Neurosci. 6, 105-122. Abstract
Franks, C. J., Pemberton, D., Vinogradova, I., Cook, A., Walker, R. J., and Holden-Dye, L. (2002). Ionic basis of the resting membrane potential and action potential in the pharyngeal muscle of Caenorhabditis elegans. J. Neurophysiol. 87, 954-961. Abstract
Fujiwara, M., Sengupta, P., and McIntire, S. L. (2002). Regulation of body size and behavioral state of C. elegans by sensory perception and the EGL-4 cGMP-dependent protein kinase. Neuron 36, 1091-1102. Abstract Article
Hibbs, R. E., and Gouaux, E. (2011). Principles of activation and permeation in an anion-selective Cys-loop receptor. Nature 474, 54-60. Abstract Article
Hobson, R. J., Hapiak, V. M., Xiao, H., Buehrer, K. L., Komuniecki, P. R., and Komuniecki, R. W. (2006). SER-7, a Caenorhabditis elegans 5-HT7-like receptor, is essential for the 5-HT stimulation of pharyngeal pumping and egg laying. Genetics 172, 159-169. Abstract Article
Horvitz, H. R., Chalfie, M., Trent, C., Sulston, J. E., and Evans, P. D. (1982). Serotonin and octopamine in the nematode Caenorhabditis elegans. Science 216, 1012-1014. Abstract Article
Husson, S. J., Clynen, E., Baggerman, G., Janssen, T., and Schoofs, L. (2006). Defective processing of neuropeptide precursors in Caenorhabditis elegans lacking proprotein convertase 2 (KPC-2/EGL-3): mutant analysis by mass spectrometry. J. Neurochem. 98, 1999-2012. Abstract Article
Husson, S. J., Janssen, T., Baggerman, G., Bogert, B., Kahn-Kirby, A. H., Ashrafi, K., and Schoofs, L. (2007). Impaired processing of FLP and NLP peptides in carboxypeptidase E (EGL-21)-deficient Caenorhabditis elegans as analyzed by mass spectrometry. J. Neurochem. 102, 246-260. Abstract Article
Ishikawa, T., Mizunoya, W., Shibakusa, T., Inoue, K., and Fushiki, T. (2006). Transforming growth factor-beta in the brain regulates fat metabolism during endurance exercise. Am. J. Physiol. Endocrinol. Metab. 291, E1151-1159. Abstract Article
Kang, C., You, Y. J., and Avery, L. (2007). Dual roles of autophagy in the survival of Caenorhabditis elegans during starvation. Genes Dev. 21, 2161-2171. Abstract Article
Kimura, K. D., Tissenbaum, H. A., Liu, Y., and Ruvkun, G. (1997). daf-2, an insulin receptor-like gene that regulates longevity and diapause in Caenorhabditis elegans. Science 277, 942-946. Abstract Article
Komatsu, H., Mori, I., Rhee, J. S., Akaike, N., and Ohshima, Y. (1996). Mutations in a cyclic nucleotide-gated channel lead to abnormal thermosensation and chemosensation in C. elegans. Neuron 17, 707-718. Abstract Article
Lee, J. S., Min, D. S., Park, C., Park, C. S., and Cho, N. J. (2001). Phytosphingosine and C2-phytoceramide induce cell death and inhibit carbachol-stimulated phospholipase D activation in Chinese hamster ovary cells expressing the Caenorhabditis elegans muscarinic acetylcholine receptor. FEBS Lett. 499, 82-86. Abstract Article
Lee, R. Y., Lobel, L., Hengartner, M., Horvitz, H. R., and Avery, L. (1997). Mutations in the alpha1 subunit of an L-type voltage-activated Ca2+ channel cause myotonia in Caenorhabditis elegans. EMBO J. 16, 6066-6076. Abstract Article
Lee, R. Y., Sawin, E. R., Chalfie, M., Horvitz, H. R., and Avery, L. (1999). EAT-4, a homolog of a mammalian sodium-dependent inorganic phosphate cotransporter, is necessary for glutamatergic neurotransmission in Caenorhabditis elegans. J. Neurosci. 19, 159-167. Abstract
Mak, H. Y., Nelson, L. S., Basson, M., Johnson, C. D., and Ruvkun, G. (2006). Polygenic control of Caenorhabditis elegans fat storage. Nat. Genet. 38, 363-368. Abstract Article
McKay, J. P., Raizen, D. M., Gottschalk, A., Schafer, W. R., and Avery, L. (2004). eat-2 and eat-18 are required for nicotinic neurotransmission in the Caenorhabditis elegans pharynx. Genetics 166, 161-169. Abstract Article
McKay, R. M., McKay, J. P., Avery, L., and Graff, J. M. (2003). C. elegans: a model for exploring the genetics of fat storage. Dev. Cell. 4, 131-142. Abstract Article
Min, D. S., Cho, N. J., Yoon, S. H., Lee, Y. H., Hahn, S. J., Lee, K. H., Kim, M. S., and Jo, Y. H. (2000). Phospholipase C, protein kinase C, Ca(2+)/calmodulin-dependent protein kinase II, and tyrosine phosphorylation are involved in carbachol-induced phospholipase D activation in Chinese hamster ovary cells expressing muscarinic acetylcholine receptor of Caenorhabditis elegans. J. Neurochem. 75, 274-281. Abstract Article
Papaioannou, S., Marsden, D., Franks, C. J., Walker, R. J., and Holden-Dye, L. (2005). Role of a FMRFamide-like family of neuropeptides in the pharyngeal nervous system of Caenorhabditis elegans. J. Neurobiol. 65, 304-319. Abstract Article
Pattillo, J. M., Yazejian, B., DiGregorio, D. A., Vergara, J. L., Grinnell, A. D., and Meriney, S. D. (2001). Contribution of presynaptic calcium-activated potassium currents to transmitter release regulation in cultured Xenopus nerve-muscle synapses. Neuroscience 102, 229-240. Abstract Article
Perez-Reyes, E. (2003). Molecular physiology of low-voltage-activated t-type calcium channels. Physiol. Rev. 83, 117-161. Abstract
Perozo, E., Cortes, D. M., and Cuello, L. G. (1999). Structural rearrangements underlying K+-channel activation gating. Science 285, 73-78. Abstract Article
Phelan, P., Bacon, J. P., Davies, J. A., Stebbings, L. A., Todman, M. G., Avery, L., Baines, R. A., Barnes, T. M., Ford, C., Hekimi, S., et al. (1998). Innexins: a family of invertebrate gap-junction proteins. Trends Genet. 14, 348-349. Abstract Article
Pocock, R., and Hobert, O. (2010). Hypoxia activates a latent circuit for processing gustatory information in C. elegans. Nat. Neurosci. 13, 610-614. Abstract Article
Posner, B. A., Gilman, A. G., and Harris, B. A. (1999). Regulators of G protein signaling 6 and 7. Purification of complexes with Gβ5 and assessment of their effects on g protein-mediated signaling pathways. J. Biol. Chem. 274, 31087-31093. Abstract Article
Raizen, D. M., and Avery, L. (1994). Electrical activity and behavior in the pharynx of Caenorhabditis elegans. Neuron 12, 483-495. Abstract Article
Raizen, D. M., Cullison, K. M., Pack, A. I., and Sundaram, M. V. (2006). A novel gain-of-function mutant of the cyclic GMP-dependent protein kinase egl-4 affects multiple physiological processes in Caenorhabditis elegans. Genetics 173, 177-187. Abstract Article
Raizen, D. M., Lee, R. Y., and Avery, L. (1995). Interacting genes required for pharyngeal excitation by motor neuron MC in Caenorhabditis elegans. Genetics 141, 1365-1382. Abstract
Ramot, D., MacInnis, B. L., Lee, H. C., and Goodman, M. B. (2008). Thermotaxis is a robust mechanism for thermoregulation in Caenorhabditis elegans nematodes. J. Neurosci. 28, 12546-12557. Abstract Article
Rand, J. B., and Nonet, M. L. (1997). Synaptic transmission. In C. elegans II, D. L. Riddle, T. Blumenthal, B. J. Meyer, and J. R. Priess, eds. (Cold Spring Harbor, Cold Spring Harbor Laboratory Press), pp. 611-643. Abstract
Reiner, D. J., and Thomas, J. H. (1995). Reversal of a muscle response to GABA during C. elegans male development. J. Neurosci. 15, 6094-6102. Abstract
Ren, P., Lim, C. S., Johnsen, R., Albert, P. S., Pilgrim, D., and Riddle, D. L. (1996). Control of C. elegans larval development by neuronal expression of a TGF-β homolog. Science 274, 1389-1391. Abstract Article
Riddle, D. L., Swanson, M. M., and Albert, P. S. (1981). Interacting genes in nematode dauer larva formation. Nature 290, 668-671. Abstract Article
Robatzek, M., Niacaris, T., Steger, K., Avery, L., and Thomas, J. H. (2001). eat-11 encodes GPB-2, a Gβ5 ortholog that interacts with Goα and Gqα to regulate C. elegans behavior. Curr. Biol. 11, 288-293. Abstract Article
Roberts, W. M., Jacobs, R. A., and Hudspeth, A. J. (1990). Colocalization of ion channels involved in frequency selectivity and synaptic transmission at presynaptic active zones of hair cells. J. Neurosci. 10, 3664-3684. Abstract
Robitaille, R., and Charlton, M. P. (1992). Presynaptic calcium signals and transmitter release are modulated by calcium-activated potassium channels. J. Neurosci. 12, 297-305. Abstract
Sanguinetti, M. C., Jiang, C., Curran, M. E., and Keating, M. T. (1995). A mechanistic link between an inherited and an acquired cardiac arrhythmia: HERG encodes the IKr potassium channel. Cell 81, 299-307. Abstract Article
Sengenes, C., Berlan, M., De Glisezinski, I., Lafontan, M., and Galitzky, J. (2000). Natriuretic peptides: a new lipolytic pathway in human adipocytes. Faseb J. 14, 1345-1351. Abstract Article
Seymour, M., Wright, K., and Doncaster, C. (1983). The action of the anterior feeding apparatus of Caenorhabditis elegans (Nematoda: Rhabditida). J. Zool. Soc. London 201, 527-539. Article
Shimozono, S., Fukano, T., Kimura, K. D., Mori, I., Kirino, Y., and Miyawaki, A. (2004). Slow Ca2+ dynamics in pharyngeal muscles in Caenorhabditis elegans during fast pumping. EMBO Rep. 5, 521-526. Abstract Article
Shtonda, B., and Avery, L. (2005). CCA-1, EGL-19 and EXP-2 currents shape action potentials in the Caenorhabditis elegans pharynx. J. Exp. Biol. 208, 2177-2190. Abstract Article
Shtonda, B. B., and Avery, L. (2006). Dietary choice behavior in Caenorhabditis elegans. J. Exp. Biol. 209, 89-102. Abstract Article
Snow, B. E., Krumins, A. M., Brothers, G. M., Lee, S. F., Wall, M. A., Chung, S., Mangion, J., Arya, S., Gilman, A. G., and Siderovski, D. P. (1998). A G protein gamma subunit-like domain shared between RGS11 and other RGS proteins specifies binding to Gbeta5 subunits. Proc. Natl. Acad. Sci. U S A 95, 13307-13312. Abstract Article
Starich, T. A., Lee, R. Y., Panzarella, C., Avery, L., and Shaw, J. E. (1996). eat-5 and unc-7 represent a multigene family in Caenorhabditis elegans involved in cell-cell coupling. J. Cell Biol. 134, 537-548. Abstract Article
Steger, K. A., and Avery, L. (2004). The GAR-3 muscarinic receptor cooperates with calcium signals to regulate muscle contraction in the Caenorhabditis elegans pharynx. Genetics 167, 633-643. Abstract Article
Steger, K. A., Shtonda, B. B., Thacker, C., Snutch, T. P., and Avery, L. (2005). The C. elegans T-type calcium channel CCA-1 boosts neuromuscular transmission. J. Exp. Biol. 208, 2191-2203. Abstract Article
Thomas, J. H. (1990). Genetic analysis of defecation in Caenorhabditis elegans. Genetics 124, 855-872.
Tsalik, E. L., Niacaris, T., Wenick, A. S., Pau, K., Avery, L., and Hobert, O. (2003). LIM homeobox gene-dependent expression of biogenic amine receptors in restricted regions of the C. elegans nervous system. Dev. Biol. 263, 81-102. Abstract Article
Tseng, G. N. (2001). I(Kr): the hERG channel. J. Mol. Cell. Cardiol. 33, 835-849. Abstract
van der Linden, A. M., Simmer, F., Cuppen, E., and Plasterk, R. H. (2001). The G-protein β-subunit GPB-2 in Caenorhabditis elegans regulates the Goα-Gqα signaling network through interactions with the regulator of G-protein signaling proteins EGL-10 and EAT-16. Genetics 158, 221-235. Abstract
Varoqui, H., Diebler, M. F., Meunier, F. M., Rand, J. B., Usdin, T. B., Bonner, T. I., Eiden, L. E., and Erickson, J. D. (1994). Cloning and expression of the vesamicol binding protein from the marine ray Torpedo. Homology with the putative vesicular acetylcholine transporter UNC-17 from Caenorhabditis elegans. FEBS Lett. 342, 97-102. Abstract Article
Vinogradova, I., Cook, A., and Holden-Dye, L. (2006). The ionic dependence of voltage-activated inward currents in the pharyngeal muscle of Caenorhabditis elegans. Invert. Neurosci. 6, 57-68. Abstract Article
Wang, Z. W., Saifee, O., Nonet, M. L., and Salkoff, L. (2001). SLO-1 potassium channels control quantal content of neurotransmitter release at the C. elegans neuromuscular junction. Neuron 32, 867-881. Abstract Article
Ward, A., Liu, J., Feng, Z., and Xu, X. Z. (2008). Light-sensitive neurons and channels mediate phototaxis in C. elegans. Nat. Neurosci. 11, 916-922. Abstract Article
*Edited by Erik M. Jorgensen. Last revised February 13, 2012. Published May 21, 2012. This chapter should be cited as: Avery, L. and You, Y.J. C. elegans feeding (May 21, 2012), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.150.1, http://www.wormbook.org.
Copyright: © 2012 Leon Avery and Young-Jai You. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
§To whom correspondence should be addressed. E-mail: lavery3@vcu.edu
 All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.
All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.