Embryo series courtesy of Einhard Schierenberg
Embryo series courtesy of Einhard SchierenbergTable of Contents
Abstract
The nematode cuticle is an extremely flexible and resilient exoskeleton that permits locomotion via attachment to muscle, confers environmental protection and allows growth by molting. It is synthesised five times, once in the embryo and subsequently at the end of each larval stage prior to molting. It is a highly structured extra-cellular matrix (ECM), composed predominantly of cross-linked collagens, additional insoluble proteins termed cuticlins, associated glycoproteins and lipids. The cuticle collagens are encoded by a large gene family that are subject to strict patterns of temporal regulation. Cuticle collagen biosynthesis involves numerous co- and post-translational modification, processing, secretion and cross-linking steps that in turn are catalysed by specific enzymes and chaperones. Mutations in individual collagen genes and their biosynthetic pathway components can result in a range of defects from abnormal morphology (dumpy and blister) to embryonic and larval death, confirming an essential role for this structure and highlighting its potential as an ECM experimental model system.
During post-embryonic development, Caenorhabditis elegans is enclosed within an exoskeleton called the cuticle (Cox et al., 1981b; Cox et al., 1981c; Kramer, 1994; Johnstone, 1994; Kramer, 1997). The cuticle is a collagenous extracellular matrix (ECM) that is synthesized by an underlying ectodermal cell layer termed the hypodermis that surrounds the body of the animal (see Figure 1). During synthesis, material is secreted from the apical membranes of the hypodermis, then polymerizes on the outer surface of the membranes where it remains in intimate contact as the mature cuticle.
Synthesis occurs five times during development, once in the embryo, then prior to molting at the end of each larval stage. Thus, with the exception of the first round of synthesis, synthesis occurs underneath an existing cuticle and requires its displacement from the membrane surface prior to, or concurrent with, secretion and polymerization of the new cuticle (see Figure 1). The old cuticle is removed by molting (Singh and Sulston, 1978).
The cuticle is a multi-functional exoskeleton. It is a highly impervious barrier between the animal and its environment. It is essential for maintenance of body morphology and integrity, and has a critical role in locomotion via attachments to body-wall muscles (Kramer et al., 1988; Von Mende et al., 1988; Johnstone et al., 1992).
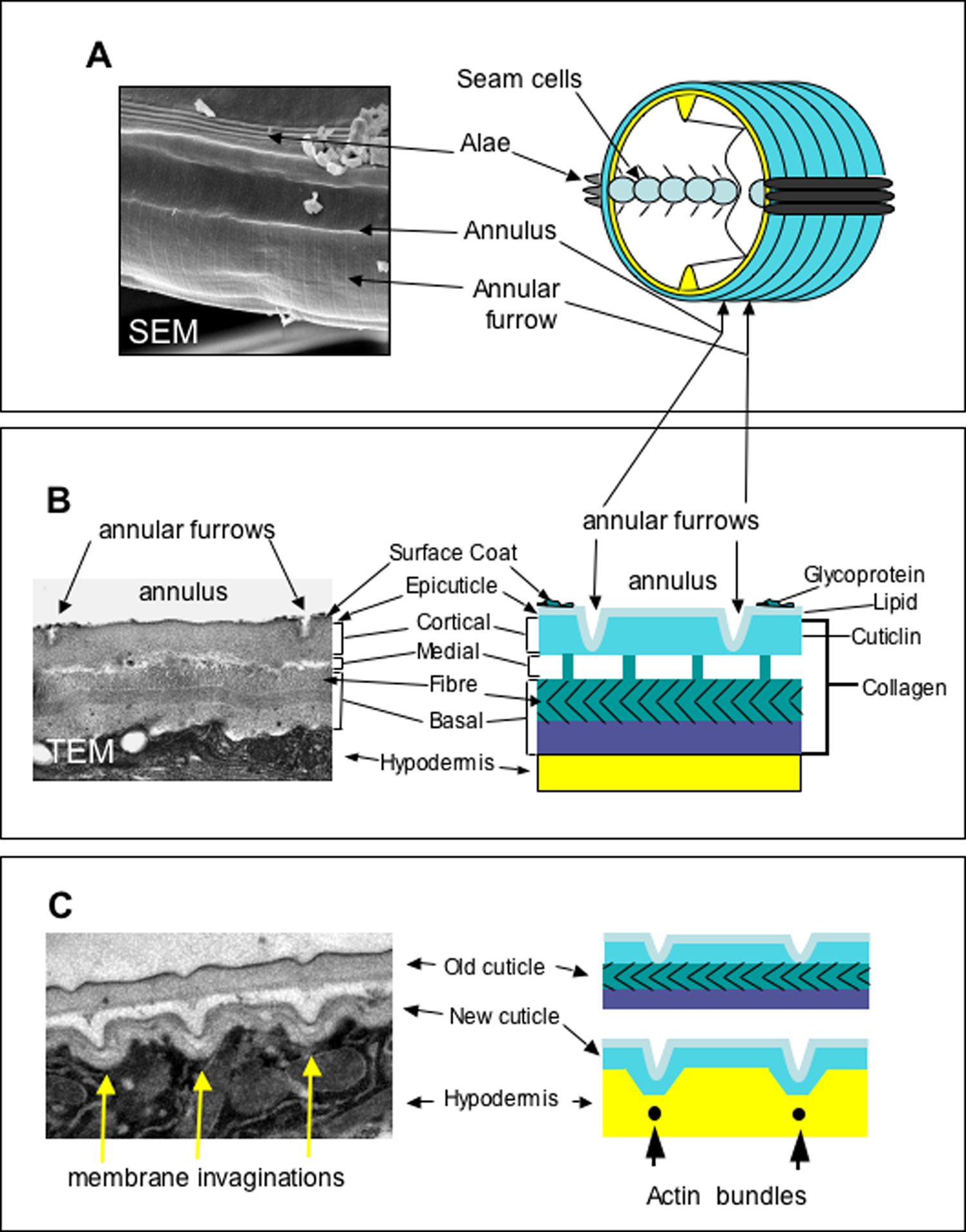
The cuticle surrounding the main body of the animal can be sub-divided into the broad dorsal and ventral regions overlying the dorsal and ventral hypodermis and the narrow lateral regions overlying the seam cells where longitudinal ridges termed alae are positioned on the cuticle of the L1 and dauer larvae and of the adult (see Figure 1A; Cox et al., 1981a; Johnstone, 2000). The alae are distinct from the rest of the cuticle both in protein composition and ultrastructure.
The dorsal and ventral cuticle has a gross multilayered ultrastructure, the detail of which varies at different developmental stages (Cox et al., 1981c; Peixoto and Desouza, 1995; Peixoto et al., 1997). There are four major layers (epicuticle, cortical, medial and basal), which can be further subdivided (see Figure 1B). The outer (cortical) layer of the dorsal and ventral regions is patterned at all developmental stages with circumferential ridges termed annuli (see Figure 1A and B). On developmental stages with alae, the annuli of the dorsal and ventral regions are separated by the alae (see Figure 1A). On stages without alae, the dorsal and ventral regions (and annuli) meet with an obvious discontinuity above the lateral seam cells. The mechanism by which the pattern of lateral alae is generated (L1 larva, dauer and adult) is unclear; however the lateral seam cells are responsible and may involve the interaction and polymerization of specific cuticulin proteins (Sapio et al., 2005). Laser ablation of individual seam cells in the L3 and early L4 results in a corresponding gap in the alae of the adult cuticle, and likewise mutants (lin-5 e1348) that lack individual seam cells also lack alae above the missing seam cells (Singh and Sulston, 1978).
During cuticle synthesis, sub-membranous actin filaments form within the hypodermal cells and are organized circumferentially around the cylindrical body of the worm (see Figure 1C), coincident with the furrows that form on the apical surface of the hypodermal cell membrane and subsequently with the furrows that delineate the boundaries of the annuli on the surface of the polymerized cuticle (see Figure 1C; Costa et al., 1997). It is therefore probable that the pattern of actin filaments has a definitive role in establishing the annular pattern on the surface of the cuticle.
|
Figure 1. The organization and structure of the C. elegans cuticle. Panel A shows a scanning electron micrograph (SEM) of the surface of a wild type adult stage animal and an accompanying schematic representation. Panel B is a transmission electron micrograph (TEM) depicting a longitudinal cross-section of the adult cuticle highlighting the distinct structural layers and their composition. With the exception of the epicuticle and surface coat, collagens are present in all major layers. Cuticlins are restricted to the cortical layer. The epicuticle contains lipids and is covered by a glycoprotein-rich coat. Panel C depicts the synthesis of a new cuticle and the associated detachment of the old cuticle. The annular pattern of the outer cuticle layer is established by invaginations in the surface of the hypodermal membrane and corresponds to the position of circumferential actin bundles which form early in lethargus. Later in lethargus, the actin bundles dissociate, the invaginations disappear and the non-invaginated inner layers of the cuticle are deposited. Thus the annular pattern is restricted to the outer layer of the cuticle.
The presence of the actin filaments and the furrows that they produce on the surface of the hypodermal membrane are transient; they are no longer present when the inner layers of the cuticle, which are not patterned by furrows, polymerize. The furrows remain in the outer layer of the cuticle, indicating that once polymerization has occurred, their presence does not require the continued presence of the cytoskeletal actin bundles. The persistence of the annular furrows in the cuticle does require the function of a specific group of cuticle collagens, discussed in detail below.
The major component of this extracellular matrix (ECM) is collagen, a protein that represents over 80% of the soluble protein released following its extraction in reducing agents. Collagens are ubiquitous structural proteins with a characteristic Glycine-X-Y tripeptide repeat, where X is frequently proline and Y hydroxyproline. The procollagens are synthesised, modified and trimerize in the endoplasmic reticulum (ER). In addition to collagens, a novel highly cross-linked insoluble class of proteins called cuticlins are associated with the cuticle. The outermost layer, the epicuticle is lipid-rich and this in turn is overlaid by a loosely associated glycoprotein-rich surface coat.
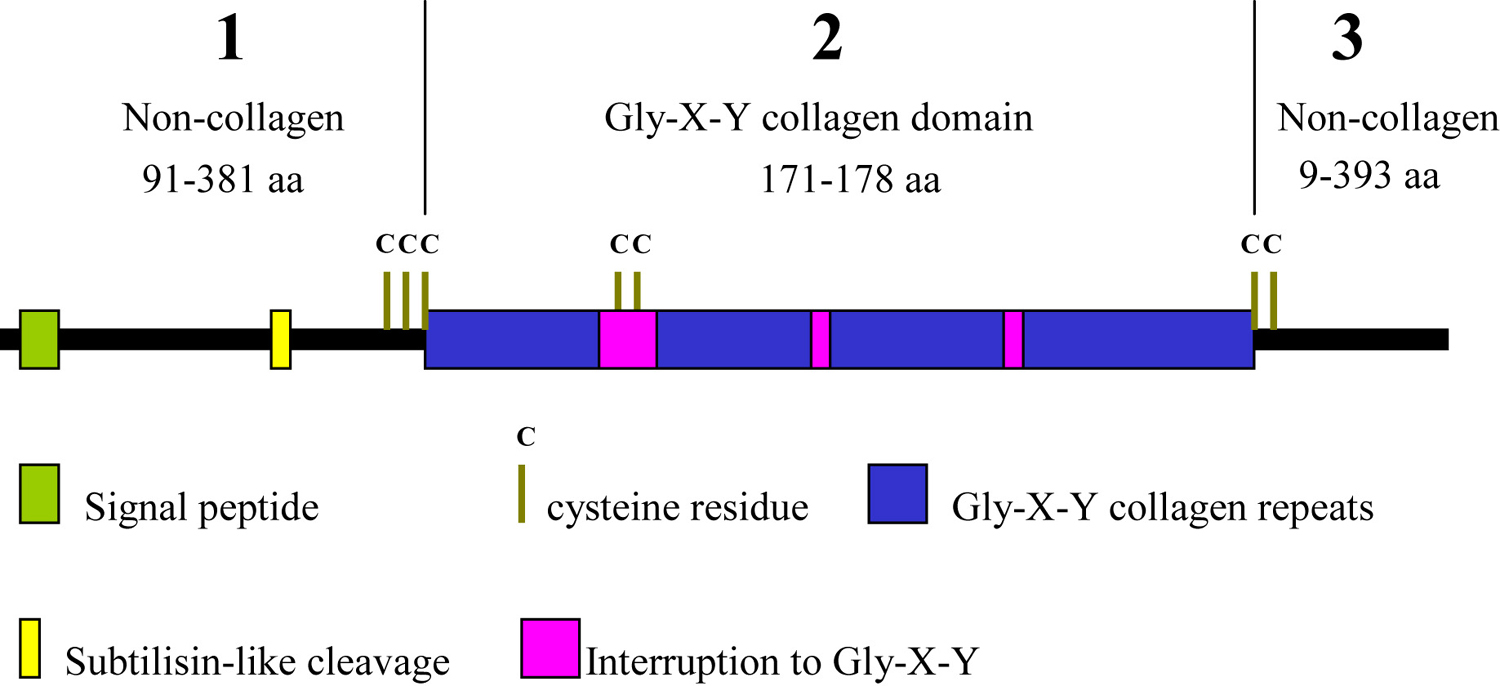
The cuticle collagens are encoded by a gene family with over 170 members (see WormBase). They have a characteristic structure (see Figure 2) of short interrupted blocks of Gly-X-Y sequence flanked by conserved cysteine residues and can be grouped into families according to homology (Johnstone, 2000). In comparison to the collagens of vertebrates, in structure and size they are most similar to the fibril-associated collagens with interrupted triple helices (FACIT).
|
Figure 2. Schematic representation of a typical cuticle collagen. The cuticle collagens share a common three-domain structure with non-collagenous amino and carboxy domains and a central Gly-X-Y collagen domain. The collagen domain contains variable small interruptions. Highly conserved clusters of cysteine residues flank the collagen domain and are present within one of the interruptions. It is predicted that proteolysis via the conserved subtilisin cleavage site removes much of the amino terminal domain of all cuticle collagens. A subset of cuticle collagens is also processed via a C-terminal BMP-like site.
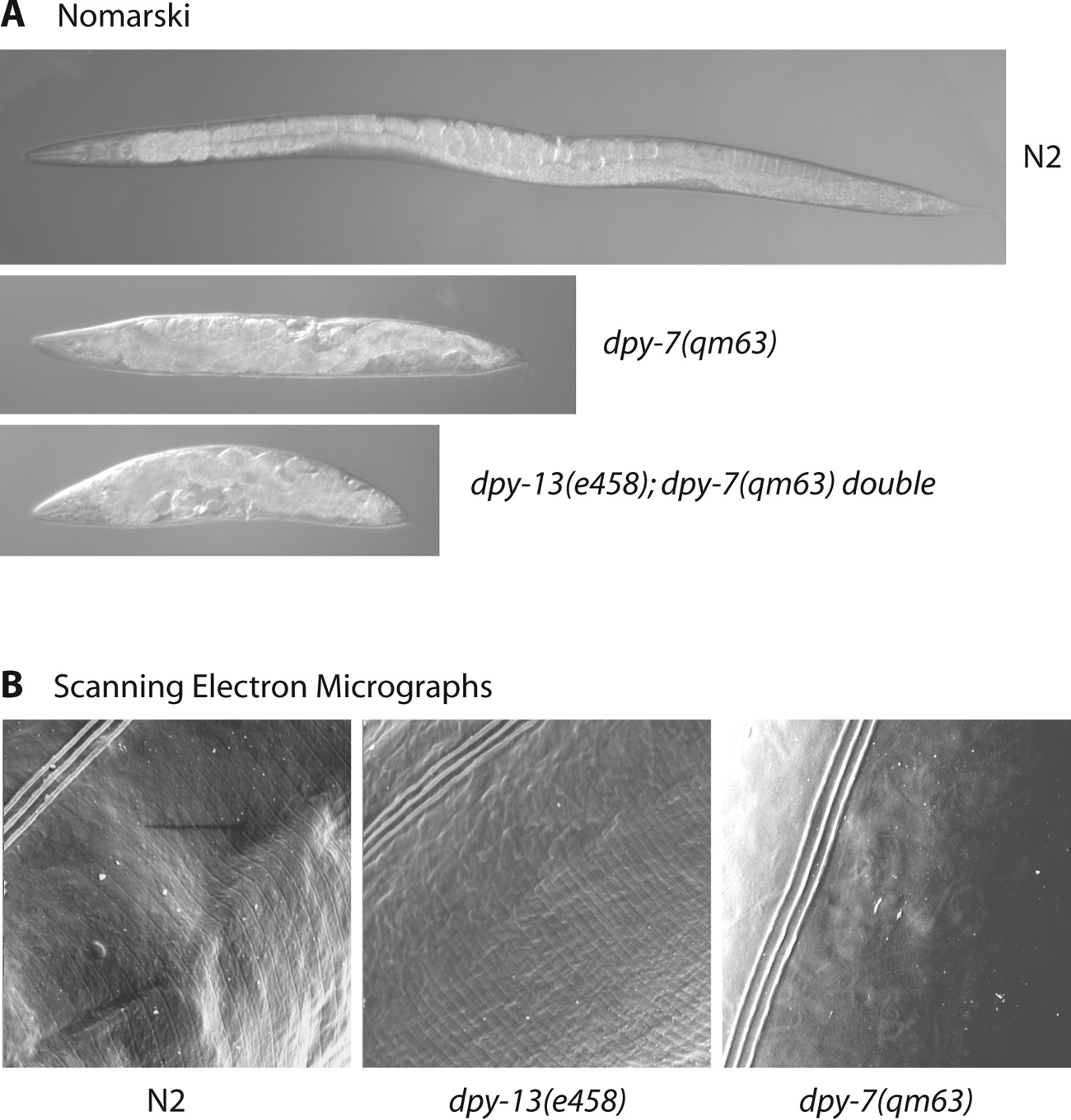
To date, mutants of 21 of the cuticle collagen genes have been identified that result in a range of informative body morphology defects including phenotypes described as, DumPY (Dpy), ROLler (Rol), BLIster (Bli), SQuaT (Sqt), Ray AbnorMal (Ram) and LONg (Lon) (see Table 1 and Figure 3). As extensive genetic screens have failed to detect mutants of the majority of the ~170 cuticle collagen genes, it is reasonable to assume that mutation of most results in phenotypes too subtle to be readily detected. Additionally, RNAi data may be confusing as there is sufficient sequence homology between many of the cuticle collagen genes that in some cases, RNAi effects could be the result of reduction of mRNA of several genes. Some cuticle collagen genes have been shown to encode components of cuticular sub-structures with specific functions. Mutants of the cuticle collagen genes dpy-2, 3, 7, 8 or 10 are dumpy (short fat worms) and have no cuticular annuli (see Figure 3B; McMahon et al., 2003). DPY-7 and DPY-10 have been shown to locate to narrow circumferential bands positioned within or beneath the annular furrows (see Figure 4). It has been proposed that DPY-2, 3 and 8 are also components of these bands. In mutants of any one of the five "no annuli" dpy genes, the DPY-7/DPY-10 containing bands are absent, and thus all five encoded collagens are believed to be obligate partners required for generation of these annular-furrow band structures. The structures are necessary for the presence/persistence of the annular furrows to which they locate, and hence the annuli that they define.
Table 1. C. elegans cuticle collagen genes that have associated phenotypes. Cosmid clone associated with the collagens genes are depicted. Groups 1, 2, 3, dpy-7 and dpy-2 refer to the structurally related collagen families based on position of conserved cysteines and Gly-X-Y interruptions (Johnstone 2000). Phenotype listed have been describe for one or more alleles of the listed genes. Dpy, Dumpy; RRol, Right-handed roller; LRol, Left-handed Roller; Lon, long; Dom, dominant; Bli, blister; wDpy, weak Dumpy; DLRol, Dumpy Left-handed roller; Ram, Ray abnormal.
| Gene | Cosmid | Group | Phenotypes |
|---|---|---|---|
| sqt-1 | B0491.2 | 1 | Dpy, RRol, LRol, Lon, Dom |
| dpy-17 | F54D8.1 | 1 | Dpy |
| lon-3 | ZK836.1 | 1 | Lon |
| rol-6 | T01B7.7 | 1 | RRol, Dpy, Dom |
| dpy-5 | F27C1.8 | 1 | Dpy |
| bli-1 | C09G5.6 | 1 | Bli |
| dpy-9 | T21D12.1 | 2 | Dpy |
| sqt-2 | C01B12.1 | 2 | wDpy/ RRol |
| rol-8 | ZK1290.3 | 2 | LRol |
| bli-2 | F59E12.12 | 2 | Bli |
| dpy-3 | EGAP7.1 | 2 | Dpy, DLRol |
| ram-3 | F38A3.1 | 2 | Ram/Dpy |
| rol-1 | Y57A10A.11 | 3 | LRol/wDpy |
| ram-4 | F36A4.10 | 3 | Ram |
| sqt-3 | F23H12.4 | 3 | Dpy, LRol, Dom |
| dpy-13 | F30B5.1 | 3 | Dpy |
| dpy-4 | Y41E3.2 | 3 | Dpy |
| dpy-7 | F46C8.6 | dpy-7 | Dpy, DLRol |
| dpy-8 | C31H2.2 | dpy-7 | Dpy, DLRol |
| dpy-2 | T14B4.7 | dpy-2 | Dpy, DLRol |
| dpy-10 | T14B4.6 | dpy-2 | Dpy, DLRol, LRol, Dom |
Conversely, annuli are present on mutants of dpy-5 or dpy-13, although they are similarly dumpy in phenotype to those with no annuli. However, the annuli are closer together (see Figure 3B). Consistent with its role in promoting the normal breadth of annuli, DPY-13 locates in a broad circumferential band within each annulus (see Figure 4). Double mutants between any member of the “no annuli” group of dpy genes and the “narrow annuli” group display a compounded severe dumpy phenotype (see Figure 3A).
|
Figure 3. Cuticle collagen mutants and phenotypes. Panel A depicts Normaski images of a wild type animal, a dpy-7(qm63) and a dpy-13(e458);dpy-7(qm63) double adult stages. The dpy-13(e458) mutant is comparably Dpy to the dpy-7(qm63) mutant (not shown) whereas the dpy-13(e458);dpy-7(qm63) double displays a more severe dumpiness than either single mutant. Panel B shows scanning electron micrographs of the surface of adult stage wild type and mutant animals. The annuli of the mutant dpy-13(e458) are narrower than those of the wild type and are completely absent from dpy-7(qm63) mutants.
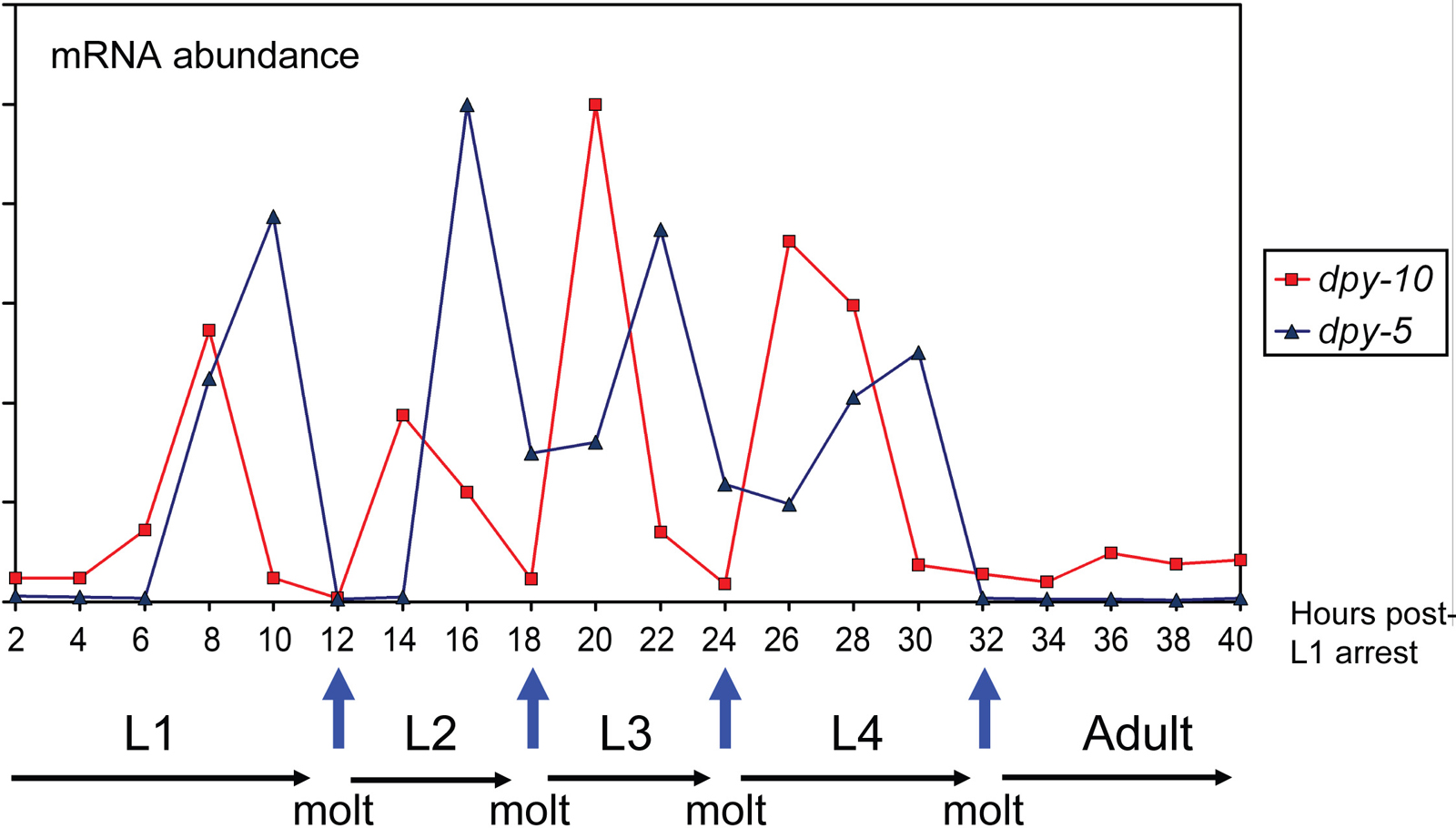
The cuticle is synthesized at the end of embryogenesis prior to hatching and then prior to molting at the end of each larval stage. During each cuticle synthetic period, the cuticle collagen genes are expressed in a distinct temporal series, the pattern of which is repeated at each synthetic period (Johnstone and Barry, 1996). According to their time of expression within this series, the cuticle collagen genes can be described as early, intermediate, or late, corresponding to peaks of mRNA abundance at approximately four hours prior, two hours prior, and concurrent with secretion of each new cuticle, respectively. A comparison of the timing of expression of the early gene dpy-10 and intermediate dpy-5 is given in Figure 5.
|
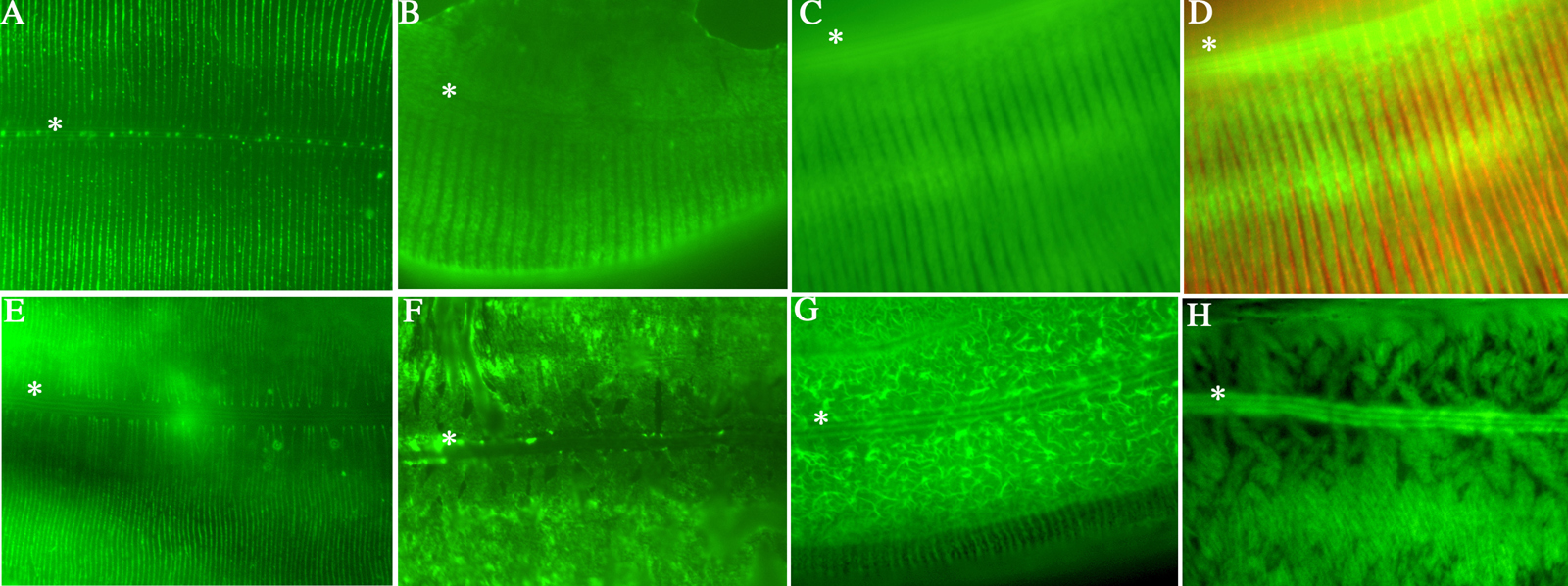
Figure 4. Localization of cuticle collagens. All images are of the adult cuticle and the position of the lateral alae is indicated by an asterix. (A) Immunolocalization of DPY-7 (McMahon et al., 2003) in the N2 cuticle; the regular stripe pattern is evident and corresponds to the location of the annular furrows. (B) Immunolocalization of epitope-tagged DPY-13 in the N2 cuticle; these broader stripes locate to annuli. (C) Localization of COL-19::GFP tagged protein (Thein et al., 2003) in a wild type background; the COL-19 localization is comparable to the DPY-13 localization depicted in B, but is also abundant in the lateral alae. (D) Immunolocalization of DPY-7 (red) and localization of COL-19::GFP (green) demonstrating that DYP-7 is expressed in the gaps between the COL-19 stripes. (E) Immunolocalization of DPY-7 in a dpy-13(e458) mutant background. Consistent with the narrower annuli of the dyp-13 mutant the DPY-7 stripes are closer together (compare to A). (F) Immunolocalization of epitope-tagged DPY-13 in the dpy-7(e88) mutant background. The regular striped pattern is lost. (G) Localization of COL-19::GFP tagged protein in a dpy-13(e458) mutant background. The COL-19 pattern is disrupted proximal to the alae, however the dorsal and ventral annular stripes are present (see bottom of images). The stripes are narrower, consistent with the narrow annuli of the dyp-13(e458) mutant (compare to wild type in C). (H) Localization of COL-19::GFP tagged protein in a in dpy-7(e88) mutant background. The regular stripe pattern is lost.
|
Figure 5. Developmental timing of collagen expression. Messenger RNA abundance of individual collagen genes display periodic fluctuations associated with cuticle synthesis.
For the two sets of dpy cuticle collagen genes discussed above, the dpy-2, dpy-3, dpy-7, dpy-8, and dpy-10 genes are all early expressed, whereas the dpy-5 and dpy-13 genes are intermediate expressed. This is consistent with a model where the individual collagen components of the same cuticle sub-structure are synthesized at the same time and different sub-structures may be synthesized at different times.
The cuticlins comprise the insoluble material that remains after extensive ionic detergent and reducing agent extraction of the cuticle. This unusual structural component is highly cross-linked, predominantly via tyrosine, a cross-link that is particularly abundant in the outermost cortical layers and in the dauer cuticle. The cuticlin proteins have cysteine-rich zona pellucida-like domains, are expressed from the hypodermal cells and play stage-specific roles in formation of the seam cell derived cuticular alae. cut-1 was the first CUTiclin gene to be identified (Sebastiano et al., 1991) and together with cut-6 is expressed in the dauer cuticle, underlying and adjacent to the dauer alae, respectively. The CUT-1 protein is expressed in the seam cell (Sebastiano et al., 1991) whereas CUT-6 is expressed at the periphery of the adjacent dorsal and ventral hypodermal cells (Muriel et al., 2003). CUT-1 and CUT-6 are involved in dauer alae formation and ultimately maintain the correct radial body morphology in this long-lived stage (Muriel et al., 2003; Sapio et al., 2005). CUT-3 is similarly associated with alae formation and body morphology in the L1 larvae, while CUT-5 is involved in L1 and dauer alae formation whereas CUT-4 is involved in the adult cuticle formation (Sapio et al., 2005). It has been hypothesized that the cuticlins, together with unidentified cuticle components, are enzymatically polymerized to constrict the seam cell-derived cuticle thereby forming the distinctive cuticular alae (Sapio et al., 2005).
Overlying the nematode cuticle is the lipid-rich epicuticle that is then covered by the glycoprotein-rich negatively charged surface coat. This labile accessory layer is synthesized from the excretory system and gland cells and has been associated with immune evasion in several parasitic nematode species (Page et al., 1992; Blaxter et al., 1992). In the natural environment it may function to aid locomotion and prevent predatory microbe adhesion to C. elegans. The Srf (SuRFace) class of mutants differ in their stage-specific expression of exposed carbohydrate epitopes (Politz et al., 1990; Politz and Philipp, 1992). Of the nine Srf mutants isolated only one has been cloned to date, namely srf-3, and represents a nucleotide sugar transporter UDP-Gal (Hoflich et al., 2004). Null mutants in this excretory cell-synthesised protein have altered surface glycoprotein expression, a weakened cuticle and display altered adherence properties to certain bacterial species (Hoflich et al., 2004). Glycoconjugate analysis of srf-3 mutants confirmed that, in comparison to wild type nematodes, N and O-linked galactose- containing glycans were significantly depleted, corroborating a role for these surface coat expressed glycans in bacterial adhesion (Cipollo et al., 2004). Several srf genes have been re-isolated in mutant screens for nematodes resistant to infection with Microbacterium nematophilium (Gravato-Nobre et al., 2005). Some of the additional Bus (Bacterially UnSwollen) mutants were also found to have weakened cuticles with unusual lectin binding properties and an associated skiddy movement phenotype (Gravato-Nobre et al., 2005). It is expected that other member of the Srf and Bus mutant class will encode components of the glycoprotein biosynthetic pathway or will be essential components of the surface coat. For additional background information regarding carbohydrates and glycosylation see the Carbohydrates and glycosylation chapter in WormBook.
The cuticle is synthesized during late embryogenesis, then shed and re-synthesized at each larval stage beneath the existing cuticle, which is subsequently shed by a process called molting (Singh and Sulston, 1978). Molting can be subdivided into lethargus, apolysis and ecdysis. Lethargus is characterised by a gradual decrease in general activity and feeding, followed by the ensuing separation of the old cuticle that defines apolysis. The nematode then rotates rapidly around its longitudinal axis, in so doing loosening the old cuticle. The shedding and emergence from the old cuticle during ecdysis completes the molting process. This complex developmental step involves extensive tissue remodelling and is assisted by numerous proteases (Davis et al., 2004; Brooks et al., 2003). The control factors involved in nematode molting and cuticle synthesis remain to be conclusively defined, however evidence does exist linking cholesterol and steroid hormones to this pathway (Yochem et al., 1999; Kuervers et al., 2003). Furthermore, the involvement of orphan nuclear hormone receptors in the process including, NHR-23 and NHR-25 is evident (Kostrouchova et al., 2001; Gissendanner and Sluder, 2000). A genome-wide RNAi-based screen has uncovered no fewer that 159 genes that are involved in the molting process; including transcription factors, secreted peptides, transmembrane and extracellular matrix proteins as well as proteases and peroxidases (Frand et al., 2005).
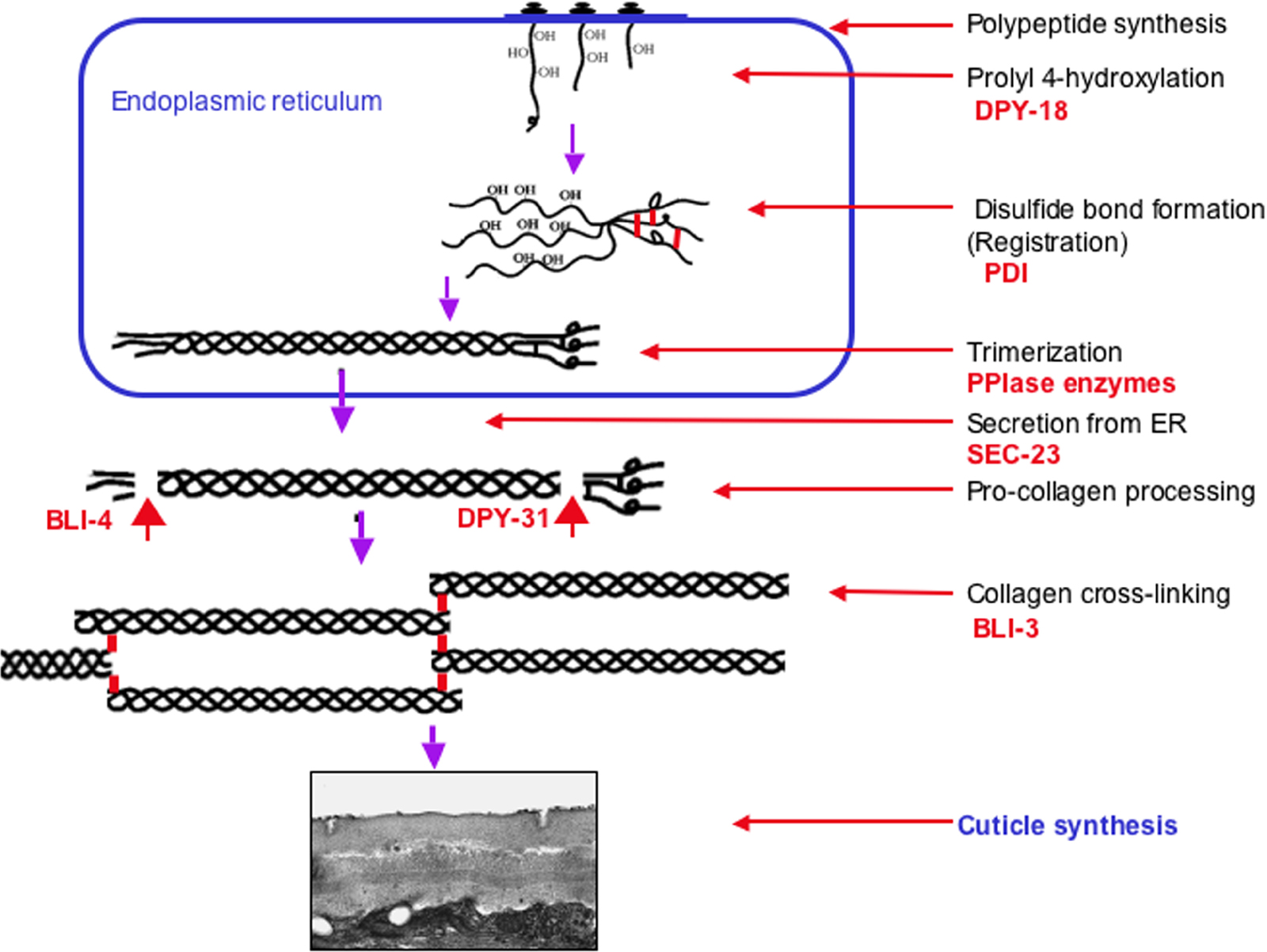
Collagen biosynthesis is a complex multi-step process in vertebrates (Prockop and Kivirikko, 1995; Myllyharju and Kivirikko, 2004). These modifications occur both intra- and extra-cellularly and involve chaperones and numerous modifying, folding and processing enzymes. It is expected that similar steps are involved in nematode collagen biosynthesis (see Figure 6) and is supported by the fact that mutations in some of these enzymes result in profound body morphology phenotypes (Page and Winter, 2003).
The first important co-translational modification of procollagen is prolyl 4-hydroxylation which allows its proper folding into a thermally-stable form (see Figure 6). The prolyl 4-hydroxylase enzymes (α-subunits) associate with the β-subunit protein disulfide isomerase (PDI) to form active, soluble ER-resident enzyme complexes. DPY-18 encodes a critical α-subunit of the multi-component enzyme prolyl 4-hydroxylase (Winter and Page, 2000; Hill et al., 2000; Friedman et al., 2000). Null mutants of dpy-18 are medium Dpy and the cuticle collagens have a corresponding reduction in their hydroxyproline content (Winter and Page, 2000; Friedman et al., 2000). The combination of dpy-18 and RNA interference of a second α-subunit phy-2 (Winter and Page, 2000) or by crossing with a phy-2 deletion mutant (Friedman et al., 2000), both result in embryonic lethality. Embryonic death was also noted following RNAi of the sole pdi-2 β-subunit-encoding gene (Winter and Page, 2000). Unique active complexes between these three subunits were detected, both in vitro and in vivo; namely, mixed DPY-18/PHY-2/(PDI-2)2 tetramers and DPY-18/PDI-2 and PHY-2/PDI-2 dimers (Myllyharju et al., 2002).
|
Figure 6. Model of cuticle collagen biogenesis. This model is based on the vertebrate fibrillar collagen assembly pathway (Prockop and Kivirikko, 1995) with adaptation specific to C. elegans cuticle collagens. Steps within the ER include; proline hydroxylation by DPY-18, disulfide bond formation and proline isomerisation, by PDI and proline isomerases respectively. Trimerization probably occurs within the ER as glycine-substitition mutants (which interfere with trimerization) cause retention of the mutant collagen within the ER. Export from the ER requires a functional COPII-pathway as loss of sec-23 causes ER retention of cuticle collagen. All cuticle collagens are believed to be N-terminally processed by the subtilisin-like protease BLI-4, whereas a subset of collagen are C-terminally processes by the BMP-like protease DPY-31. The final step in collagen maturation involves tyrosine crosslink formation and is catalized by peroxidase enzymes including BLI-3.
The next important step in vertebrate collagen folding, that precedes trimer formation is the correct registration of the monomers. Collagen trimerization can be initiated from either N- or C-terminal directions, and may be set up via PDI catalysed disulfide bond formation. The ER-resident enzyme PDI (see Figure 7) may therefore play multiple independent roles in the proper biosynthesis of collagen (see Figure 6).
|
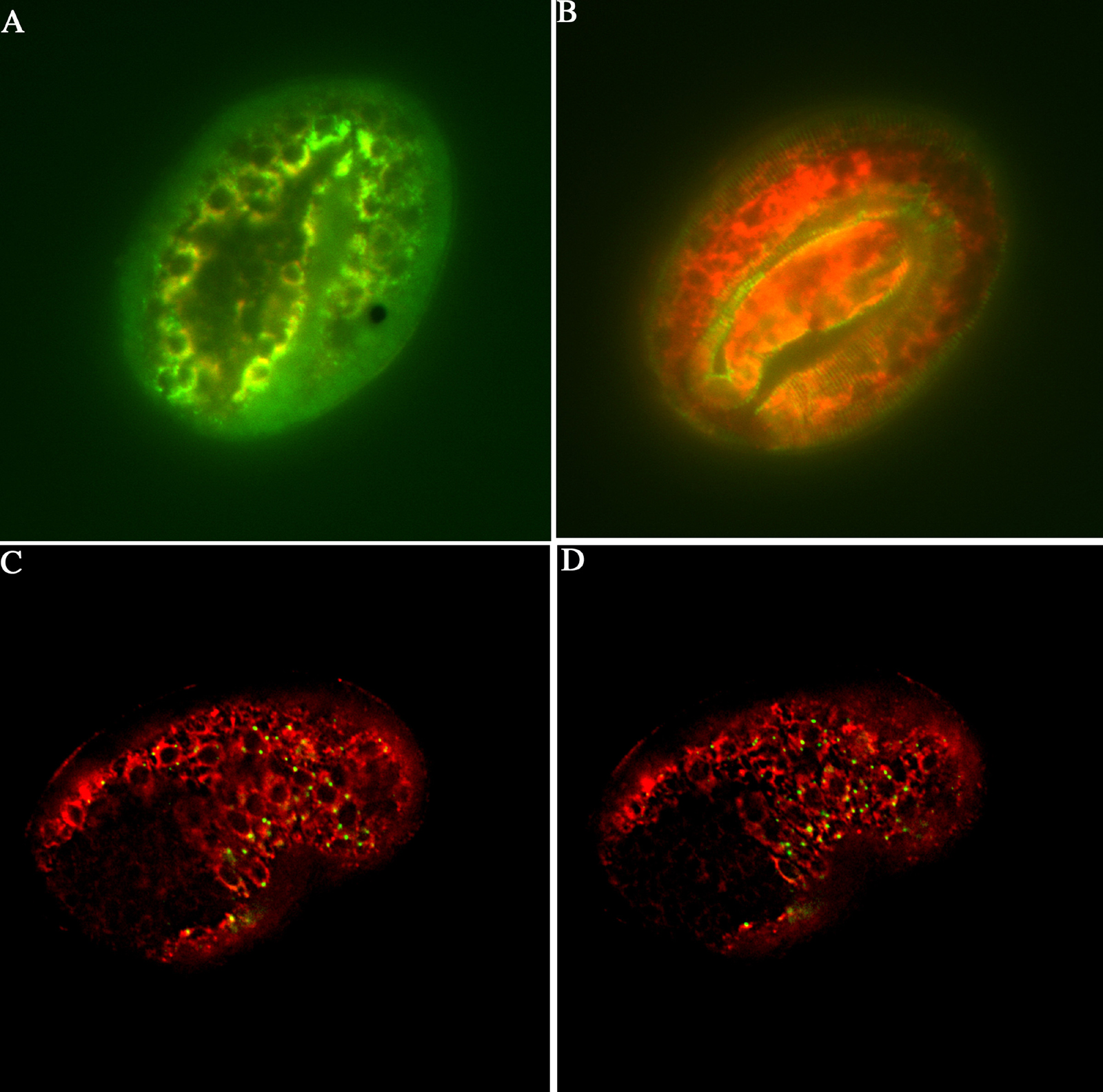
Figure 7. Subcellular localization of SEC-23 and PDI-2 in C. elegans. (A) DPY-7 (green) and PDI-2 (red) double staining of a 1.5-fold embryo. At this stage both PDI-2 and DPY-7 are co-localised in the ER (yellow). (B) DPY-7 (green) and PDI-2 (red) double staining of an elongated embryo. At this stage DPY-7 (green) has been secreted and is present in the newly formed cuticle. PDI-2 (red) remains resident in the ER. (C and D) PDI-2 (red) and SEC-23::GFP (green) localisation in early comma stage embryos. Two images of the same embryo taken at focal planes 1 μm apart. SEC-23 localises to foci on the surface of the ER as defined by the ER marker PDI-2.
The C. elegans collagens have highly conserved C-terminal cysteine clusters that may be involved in such registration events. The trimerization of imino-rich (25% proline and hydroxyproline) collagen is rate-limited by the slow cis-trans proline isomerization, and requires the assistance of peptidyl prolyl cis-trans isomerase (PPIase) enzymes of the CYclophiliN (CYN) and FK506 Binding protein (FKB) class (Bachinger, 1987; Steinmann et al., 1991). These genes constitute large, presumably redundant, families in the worm including several ER-resident isoforms that may perform this function (Page et al., 1996; Bell et al., 2006).
SEC-23 is an integral component of COPII secretory vesicles involved in transport from the ER to golgi apparatus. Loss of sec-23 in C. elegans has been shown to cause accumulation of cuticle collagen in the ER suggesting that cuticle collagen secretion occurs through the COPII pathway (Roberts et al., 2003).
Prior to collagen multimerization the procollagen trimers must first be processed to remove the N- and C-terminal teleopeptides. These catalytic events alter protein solubility, permit cross-linking and ultimately lead to the formation of the final insoluble matrix (see Figure 6). The importance of both N-terminal processing and C-terminal cross-linking has been demonstrated in the cuticle collagen SQT-1. RRol phenotypes affecting the subtilisin-like processing site result from a failure to process this site whereas LRol phenotypes, associated with the C-terminal conserved cysteine, interfere with non-reducible cross-linking of this collagen (Yang and Kramer, 1999).
The Kex2, subtilisin-like serine protease encoding gene bli-4 is essential for post-embryonic viability in C. elegans (Peters et al., 1991). All cuticle collagens possess a highly conserved N-terminal cleavage site for BLI-4, a site that has been experimentally demonstrated to be essential for the proper processing of the collagens ROL-6 and SQT-1 (Yang and Kramer, 1994). Null mutants of bli-4 are embryonically lethal, whereas partial loss-of-function mutants were viable but Bli and Dpy (Peters et al., 1991; Thacker et al., 1995). Furthermore, RNAi screens targeting bli-4 also described an associated molt defect (Kamath et al., 2003).
The C-terminal processing of vertebrate fibrillar collagens is an essential step and is carried out by zinc metalloproteases of the astacin, bone morphogenic protein (BMP) class (Canty and Kadler, 2005; Pappano et al., 2003). The majority of nematode collagens have very short C-terminal non Gly-X-Y domains and until recently this processing step was not considered to be important for their maturation. Isolation and characterization of mutations in the BMP encoding gene dpy-31 result in temperature sensitive severe Dpy, Emb and Let phenotypes in the worm, demonstrating that dpy-31 encodes an essential procollagen C-peptidase (Novelli et al., 2004; Figure 6). This enzyme is expressed in the hypodermis and is required for normal cuticle collagen secretion. Extragenic suppressors of dpy-31 include alleles of the cuticle collagen genes sqt-3 and dpy-17 (Novelli et al., 2006). Several sqt-3 suppressors are themselves cold-sensitive lethal, which is a direct contrast to sqt-3 and dpy-31 nulls, which are temperature sensitive lethal (Novelli et al., 2006). The C-terminal non Gly-X-Y domain of SQT-3 is a major substrate for this enzyme, the predicted cleavage site being adjacent to the C-terminal tyrosine cross-linking residues (Novelli et al., 2004; Novelli et al., 2006).
The final steps in collagen maturation and cuticle synthesis involve the structural cross-linking events. The cuticle collagens and cuticlins are covalently cross-linked via unusual di- and tri- tryrosine cross-links. These non-reducible cross-links impart the characteristic strength and integrity to the cuticle (Page, 2001 ) and differ from the hydroxylysine-derived cross-links of the vertebrate collagens (Myllyharju and Kivirikko, 2001). The major cuticle cross-linking enzyme is a dual oxidase enzyme encoded by the bli-3 locus (Edens et al., 2001; Simmer et al., 2003; Figure 6). This large enzyme has a signal peptide, a membrane-bound peroxide generating domain and a functionally-active peroxidase domain (Edens et al., 2001). It is proposed that the BLI-3 enzyme functions in a coupled reaction with the NADPH oxidase portion generating the hydrogen peroxide for the cross-linking reaction, which is then catalysed by the peroxidase portion (Edens et al., 2001). The peroxidase domain does however lack residues considered critical for haem-binding and therefore cannot be considered to be "classical" peroxidases. RNAi (Edens et al., 2001) or single point mutations (Simmer et al., 2003) in this enzyme result in worms with weakened cuticles that lack tyrosine cross-links. The resulting adult worms are devoid of cuticle struts and exhibit Dpy and Bli phenotypes. This result is consistent with the BLI-3 enzyme having the proposed dual function, but also supports a sole function in generation of hydrogen peroxide that is utilized by an as-yet unidentified Haem peroxidase.
Collagen biogenesis and ECM function play a critical role in organogenesis and body morphology in all metazoans. Defects in these processes result in a variety of debilitating conditions of humans, including osteogenesis imperfecta, Ehlers-Danlos syndrome, Epidermolysis bullosa, several chondrodysplasias and excessive fibrosis associated with wound healing and liver disease. Additionally, the capacity to control and engineer ECM synthesis will play a critical role in any future attempts at organ culture. Although the C. elegans cuticle is a highly specialized ECM with distinct and complex features, its biogenesis involves molecules, mechanisms and pathways that are shared with vertebrates. The extreme genetic tractability makes it an excellent experimental system for the dissection of ECM formation.
Bachinger, H.P. (1987). The influence of peptidyl-prolyl cis-trans isomerase on the in vitro folding of type III collagens. J. Biol. Chem. 262, 17144–17148. Abstract
Bell, A., Monaghan, P., and Page, A.P. (2006). Peptidyl-prolyl cis-trans isomerases (immunophilins) and their roles in parasite biochemistry, host-parasite interaction and antiparasitic drug action. Int. J. Parasitol. 36, 261–276. Abstract Article
Blaxter, M.L., Page, A.P., Rudin, W., and Maizels, R.M. (1992). Nematode surface coats: actively evading immunity. Parasitol. Today 8, 243–247. Abstract Article
Brooks, D.R., Appleford, P.J., Murray, L., and Isaac, R.E. (2003). An essential role in molting and morphogenesis of Caenorhabditis elegans for ACN-1, a novel member of the angiotensin-converting enzyme family that lacks a metallopeptidase active site. J. Biol. Chem. 278, 52340–52346. Abstract Article
Canty, E.G., and Kadler, K.E. (2005). Procollagen trafficking, processing and fibrillogenesis. J. Cell. Sci. 118, 1341–1353. Abstract Article
Cipollo, J.F. Awad, A.M., Costello, C.E. and Hirschberg, C.B. (2004). srf-3, a mutant of Caenorhabditis elegans, resistant to bacterial infection and to biofilm binding, is deficient in glycoconjugates. J. Biol. Chem. 279, 52893–52903. Abstract Article
Costa, M., Draper, B.W., and Priess, J.R. (1997). The role of actin filaments in patterning the Caenorhabditis elegans cuticle. Dev. Biol. 184, 373–384. Abstract Article
Cox, G.N., Kusch, M., DeNevi, K., and Edgar, R.S. (1981a). Temporal regulation of cuticle synthesis during development of Caenorhabditis elegans. Dev. Biol. 84, 277–285. Article
Cox, G.N., Kusch, M., and Edgar, R.S. (1981b). Cuticle of Caenorhabditis elegans: its isolation and partial characterisation. J. Cell Biol. 90, 7–17. Abstract Article
Cox, G.N., Straprans, S., and Edgar, R.S. (1981c). The cuticle of Caenorhabditis elegans. II Stage-specific changes in ultrastructure and protein composition during postembryonic development. Dev. Biol. 86, 456–470. Abstract Article
Davis, M.W., Birnie, A.J., Chan, A.C., Page, A.P., and Jorgensen, E.M. (2004). A conserved metalloprotease mediates ecdysis in Caenorhabditis elegans. Development 131, 6001–6008. Abstract Article
Edens, W.A., Sharling, L., Cheng, G., Shapira, R., Kinkade, J.M., Lee, T., Edens, H.A., Tang, X., Sullards, C., Flaherty, D.B., et al. (2001). Tyrosine cross-linking of extracellular matrix is catalysed by Duox, a multidomain oxidase/peroxidase with homology to the phagocyte oxidase subunit gp91/phox. J. Cell Biol. 154, 879–891. Abstract Article
Frand, A.R., Russel, S., and Ruvkun, G. (2005). Functional genomic analysis of C. elegans molting. PLoS Biol. 3, 1719–1733. Abstract Article
Friedman, L., Higgin, J.J., Moulder, G., Barstead, R., Raines, R.T., and Kimble, J. (2000). Prolyl 4-hydroxylase is required for viability and morphogenesis in Caenorhabditis elegans. Proc. Natl. Acad. Sci. U.S.A. 97, 4736–4741. Abstract Article
Gissendanner, C.R., and Sluder, A.E. (2000). Nhr-25, the Caenorhabditis elegans ortholog of ftz-f1, is required for epidermal and somatic gonad development. Dev. Biol. 221, 259–272. Abstract Article
Gravato-Nobre, M.J., Nicholas, H.R., Nijland, R., O'Rourke, D., Whittington, D.E., Yook, K.J., and Hodgkin, J. (2005). Multiple genes affect sensitivity of Caenorhabditis elegans to the bacterial pathogen Microbacterium nematophilum. Genetics 171, 1033–1045. Abstract Article
Hill, K.L., Harfe, B.D., Dobbins, C.A., and Hernault, S.W.L. (2000). dpy-18 encodes an a-subunit of prolyl 4-hydroxylase in Caenorhabditis elegans. Genetics 155, 1139–1148. Abstract
Hoflich, J., Berninsone, P., Gobel, C., Gravato-Nobre, M.J., Libby, B.J., Darby, C., Politz, S.M., Hodgkin, J., Hirschberg, C.B., and Baumeister, R. (2004). Loss of srf-3-encoded nucleotide sugar transporter activity in Caenorhabditis elegans alters surface antigenicity and prevents bacterial adherence. J. Biol. Chem. 279, 30440–30448. Abstract Article
Johnstone, I.L. (1994). The cuticle of the nematode Caenorhabditis elegans- A complex collagen structure. Bioessays 16, 171–178. Abstract Article
Johnstone, I.L. (2000). Cuticle collagen genes expression in Caenorhabditis elegans. Trends Genet. 16, 21–27. Abstract Article
Johnstone, I.L., and Barry, J.D. (1996). Temporal reiteration of a precise gene expression pattern during nematode development. EMBO J. 15, 3633–3639. Abstract
Johnstone, I.L., Shafi, Y., and Barry, J.D. (1992). Molecular analysis of mutations in the Caenorhabditis elegans collagen gene dpy-7. EMBO J. 11, 3857–3863. Abstract
Kamath, R.S., Fraser, A.G., Dong, Y., Poulin, G., Durbin, R., Gotta, M., Kanapin, A., Le Bot, N., Moreno, S., Sohrmann, M., et al. (2003). Systematic functional analysis of the Caenorhabditis elegans genome using RNAi. Nature 421, 231–237. Abstract Article
Kostrouchova, M., Krause, M., Kostrouch, Z., and Rall, J.E. (2001). Nuclear hormone receptor CHR3 is a critical regulator of all four larval molts of the nematode Caenorhabditis elegans. Proc. Natl. Acad. Sci. U.S.A. 98, 7360–7365. Abstract Article
Kramer, J.M., Johnson, J.J., Edgar, R., Basch, C., and Roberts, S. (1988). The sqt-1 gene of C. elegans encodes a collagen critical for organismal morphogenesis. Cell 55, 555–565. Abstract Article
Kramer, J.M. (1994). Structures and functions of collagens in Caenorhabditis elegans. FASEB J. 8, 329–336. Abstract
Kramer, J.M. (1997). Extracellular Matrix. In C. elegans II, D.L. Riddle, ed. (Cold Spring Harbor: Cold Spring Harbor Laboratory Press), pp. 471–500.
Kuervers, L.M., Jones, C.L., O'Neil, N.J., and Baillie, D.L. (2003). The sterol modifying enzyme LET-767 is essential for growth, reproduction and development in Caenorhabditis elegans. Mol. Genet. Genomics 270, 121–131. Abstract Article
McMahon, L., Muriel, J.M., Roberts, B., Quinn, M., and Johnstone, I.L. (2003). Two sets of interacting collagens form functionally distinct substructures within a Caenorhabditis elegans extracellular matrix. Mol. Biol. Cell 14, 1366–1378. Abstract Article
Muriel, J.M., Brannan, M., Taylor, K., Johnstone, I.L., Lithgow, G.J., and Tuckwell, D. (2003). M142.2 (cut-6), a novel Caenorhabditis elegans matrix gene important for dauer body shape. Dev. Biol. 260, 339–351. Abstract Article
Myllyharju, J., and Kivirikko, K. (2001). Collagens and collagen-related diseases. Ann. Med. 33, 7–21. Abstract
Myllyharju, J., and Kivirikko, K.I. (2004). Collagens, modifying enzymes and their mutations in humans, flies and worms. Trends Genet. 20, 33–43. Abstract Article
Myllyharju, J., Kukkola, L., Winter, A.D., and Page, A.P. (2002). The exoskeleton collagens in Caenorhabditis elegans are modified by prolyl 4-hydroxylases with unique combinations of subunits. J. Biol. Chem. 277, 29187–29196. Abstract Article
Novelli, J., Ahmed, S., and Hodgkin, J. (2004). Gene interactions in Caenorhabditis elegans define DPY-31 as a candidate procollagen C-proteinase and SQT-3/ROL-4 as its predicted major target. Genetics 168, 1259–1273. Abstract Article
Novelli, J., Page, A.P., and Hodgkin, J. (2006). The C terminus of collagen SQT-3 has complex and essential functions in nematode collagen assembly. Genetics 172, 2253–2267. Abstract Article
Page, A.P. (2001). The nematode cuticle: synthesis, modification and mutants. In, Parasitic Nematodes. Kennedy, M.W. and Harnett, W., eds., (St Albans, UK: CABI), 167–193.
Page, A.P., Hamilton, A.J., and Maizels, R.M. (1992). Toxocara canis: monoclonal antibodies to carbohydrate epitopes of secreted (TES) antigens localize to different secretion-related structures in infective larvae. Exp. Parasitol. 75, 56–71. Abstract Article
Page, A.P., MacNiven, K., and Hengartner, M.O. (1996). Cloning and biochemical characterisation of the cyclophilin homologues from the free-living nematode Caenorhabditis elegans. Biochem. J. 317, 179–185. Abstract
Page, A.P., and Winter, A.D. (2003). Enzymes involved in the biogenesis of the nematode cuticle. Adv. Parasitol. 53, 85–148. Abstract
Pappano, W.N., Steiglitz, B.M., Scott, I.C., Keene, D.R., and Greenspan, D.S. (2003). Use of BMP1/Tll1 doubly homozygous null mice and proteomics to identify and validate in vivo substrates of bone morphogenetic protein 1/tolloid-like metalloproteinases. Mol. Cell. Biol. 23, 4428–4438. Abstract Article
Peixoto, C.A., and Desouza, W. (1995). Freeze-fracture and deep-etched view of the cuticle of Caenorhabditis elegans. Tissue Cell 27, 561–568. Abstract Article
Peixoto, C.A., Kramer, J.M., and deSouza, W. (1997). Caenorhabditis elegans cuticle: A description of new elements of the fibrous layer. J. Parasitol. 83, 368–372. Abstract Article
Peters, K., McDowall, J., and Rose, A. (1991). Mutations in the bli-4(i) locus of Caenorhabditis elegans disrupt both adult cuticle and early larval development. Genetics 129, 95–102. Abstract
Politz, S.M., and Philipp, M. (1992). Caenorhabditis elegans as a model for parasitic nematodes: A focus on the cuticle. Parasitol. Today 8, 6–12. Abstract Article
Politz, S.M., Philipp, M., Estevez, M., Obrien, P.J., and Chin, K.J. (1990). Genes that can be mutated to unmask hidden antigenic determinants in the cuticle of the nematode Caenorhabditis elegans. Proc. Natl. Acad. Sci. U.S.A. 87, 2901–2905. Abstract Article
Prockop, D.J., and Kivirikko, K.I. (1995). Collagens: Molecular biology, diseases, and potential for therapy. Annu. Rev. Biochem. 64, 403–434. Abstract Article
Roberts, A., Clucas, C., and Johnstone, I.L. (2003). Loss of SEC-23 in Caenorhabditis elegans causes defects in oogenesis, morphogenesis, and extracellular matrix secretion. Mol. Biol. Cell 14, 4414–4426. Abstract Article
Sapio, M.R., Hilliard, M.A., Cermola, M., Favre, R., and Bazzicalupo, P. (2005). The zona pellucida domain containing proteins, CUT-1, CUT-3 and CUT-5, play essential roles in the development of the larval alae in Caenorhabditis elegans. Dev. Biol. 282, 231–245. Abstract Article
Sebastiano, M., Lassandro, F., and Bazzicalupo, P. (1991). Cut-1 a Caenorhabditis-elegans gene coding for a dauer-specific noncollagenous component of the cuticle. Dev. Biol. 146, 519–530. Abstract Article
Simmer, F., Moorman, C., van der Linden, A.M., Kuijk, E., van den Berghe, P.V.E., Kamath, R.S., Fraser, A.G., Ahringer, J., and Plasterk, R.H.A. (2003). Genome-wide RNAi of C. elegans using the hypersensitive rrf-3 strain reveals novel gene functions. PLoS Biol. 1, 77–84. Abstract Article
Singh, R., and Sulston, J. (1978). Some observations on moulting in Caenorhabditis elegans. Nematologica 24, 63–71.
Steinmann, B., Bruckner, P., and Superti-Furga, A. (1991). Cyclosporin A slows collagen triple-helix formation in vivo: Indirect evidence for a physiological role of peptidyl prolyl cis-trans isomerase. J. Biol. Chem. 266, 1299–1303. Abstract
Thacker, C., Peters, K., Srayko, M., and Rose, A.M. (1995). The bli-4 locus of Caenorhabditis elegans encodes structurally distinct kex2/subtilisin-like endoproteases essential for early development and adult morphology. Genes Dev. 9, 956–971. Abstract
Thein, M.C., McCormack, G., Winter, A.D., Johnstone, I.L., Shoemaker, C.B. and Page, A.P. (2003). Caenorhabditis elegans exoskeleton collagen COL-19: an adult-specific marker for collagen modification and assembly, and the analysis of organismal morphology. Dev. Dyn. 226, 523–39. Abstract Article
Von Mende, N., Bird, D., Albert, P., and Riddle, D. (1988). DPY-13: A nematode collagen gene that affects body shape. Cell 55, 567–576. Abstract Article
Winter, A.D., and Page, A.P. (2000). Prolyl 4-hydroxylase is an essential procollagen-modifying enzyme required for exoskeleton formation and the maintenance of body shape in the nematode Caenorhabditis elegans. Mol. Cell. Biol. 20, 4084–4093. Abstract Article
Yang, J., and Kramer, J.M. (1994). In vitro mutagenesis of Caenorhabditis elegans cuticle collagens identifies a potential subtilisin-like protease cleavage site and demonstrates that carboxyl domain disulfide bonding is required for normal function but not assembly. Mol. Cell. Biol. 14, 2722–2730. Abstract
Yang, J., and Kramer, J.M. (1999). Proteolytic processing of Caenorhabditis elegans SQT-1 cuticle collagen is inhibited in right roller mutants whereas cross-linking is inhibited in left roller mutants. J. Biol. Chem. 274, 32744–32749. Abstract Article
Yochem, J., Tuck, S., Greenwald, I., and Han, M. (1999). A gp330/megalin-related protein is required in the major epidermis of Caenorhabditis elegans for completion of molting. Development 126, 597–606. Abstract
*Edited by James M. Kramer and Donald G. Moerman. Last revised January 23, 2007. Published March 19, 2007. This chapter should be cited as: Page, A.P. and Johnstone, I.L. The cuticle (March 19, 2007), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.138.1, http://www.wormbook.org.
Copyright:© 2007 Antony P. Page and Iain L. Johnstone. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
§To whom correspondence should be addressed. E-mail: a.page@vet.gla.ac.uk or i.johnstone@bio.gla.ac.uk
 All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.
All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.